|
†
WYBIERZ RÓD lub GENEALOGIĘ
GENETYCZNĄ:
.
|
Strony genealogii genetycznej na
naszej witrynie (w trakcie poprawek)
1. Genealogia Y-DNA, mtDNA
i autosomów DNA. Jak zbadać swoje DNA?
2. Haplogrupy
R1i R1b euroazjatyckie i indoeuropejskie
3.
Mitochondrialne mtDNA
4.
Y-Adam, praojciec wszystkich współczesnych ludzi
5.
Geograficzna i
etniczna kolebka współczesnych ludzi
5.
Fundusz Rozwoju Rodu A00 w Kamerunie
[ 1 ]
[ 2 ]
[
3 ]
6.
Pochodzenie ludności Europy, Praindoeuropejczyków, Slowian i
Polaków
7.
Indoeuropejska geneza Scytów, Tocharów, indo-irańskich Ariów,
Anatolijczyków i Ormian)
8.
Nieindoeuropejskie i zindoeuropeizowane rody Y-DNA w Europie
9.
Praindoeuropejska geneza Słowian
10.
Polacy,
skąd i kim jesteście? Biologiczne i kulturowe korzenie
Polaków
11.
Pochodzenie ludności w dorzeczu Dunajca i jego
regionie
12.
Inne wybrane regiony
13.
"Stara Karpacka" gałąź rodu
R1a: YP343.
14.
"Polskie
Mykeny nad Dunajcem" - kultura Otomani-Füzesabony
15.
Kreacjonizm ewolucjonistyczny
|
Mój
pogląd na świat:
Kreacjonizm
ewolucjonistyczny
(the evolutionistic
creationism)
Ewolucjonizm jest czymś więcej niż tylko teorią [...].
Doktryna wiary niezmiennie
jednak głosi,
że ludzka duchowa dusza
została stworzona bezpośrednio przez Boga (św. Jan Paweł II pp, 1986)
Niezmierzony i cudowny
wszechświat
wraz z człowiekiem nie może być
dziełem przypadku
lub konieczności
(K Darwin, O powstaniu gatunków 515) |

MOJE DNA
od Y-Adama:
L74
od ojca:
R1a YP380
od
matki:
H14a |
F-b
EUREKA!
Dzięki
genealogii genetycznej już wiem, skąd pochodzą
Polacy, Słowianie,
Europejczycy, inne ludy i ja sam też!
Zobacz objaśnienia pojęć genealogii genomicznej
www.tropie.tarnow.opoka.org.pl/Genealogia_genomiczna.pdf
UWAGA: Strona w trakcie
aktualizacji!
(Uwaga, tekst tej strony, w miarę
pojawiania sie nowych odkryć i ich opracowań, bywa często
poprawiany i uzupełniany;
mogą więc pojawić się tu tymczasowe niekonsekwencje i
sprzeczności - rzeczowe i interpretacyjne; proszę o współpracę).
Jesteśmy
tu na portalu saktuarium św. Świerada i Benedykta, Pustelników
Święci nie ukrywają
swojego pochodzenia (tylko gangsterzy muszą to robić).
Pracując nad zagadnieniem pochodzenia naszych pustelników nie
można było nie zapytać o ich środowisko etniczne, a w końcu
także i genetyczne. W czasie przyjścia na świat św.
Świerada i jego pobytu nad Dunajcem
nie było tu jeszcze państwowości polskiej. Bolesław Chrobry, jak się wydaje,
zagarnął tę ziemię dopiero w 1017/1018 roku, w drodze powrotnej z Węgier po pokojowym
układzie ze św. Stefanem, a przed wyprawą kijowską. Zapewne wtedy, zwyczajem
władców państwa Polan, ostatecznie zniszczył umocnienia starodawnego grodu w Naszacowicach i nowszego w Podegrodziu (Grobla), by na to miejsce zbudować tam
swój własny symbol władzy, zwany dziś Zamczysko. Więc politycznie zorganizowana
polskość dopiero tu wkraczała. A co było wcześniej?
Czy ów naddunajecki Pustelnik był etnicznie i z krwi Polakiem i
Słowianinem? Kazano nam w to wątpić, skoro wcześniej przebywali tu, jak w całej
Polsce południowej i w Kotlinie Sądeckiej jacyś ludzie tzw. kultury przeworskiej,
wysoko zorganizowanej i podobno etnicznie celtyckiej i germańskiej (Wandalowie?). Niby
pod koniec IV wieku odeszli oni na południe, nad Dunaj i dalej, na południe i zachód, jak udowadnia archeologia
oraz południowoeuropejskie świadectwa pisane, ale jednak jakaś ludność tu
pozostała, skoro przekazała Polakom wiele starożytnych nazw rzecznych, raczej
jakoby niepolskiego, bo praindoeuropejskiego pochodzenia. Kto? Jacyś Ilirowie (Trakowie, Dakowie...) z czasów jeszcze przed-Chrystusowych?
A może o etniczności św. Świerada rozstrzyga słowiański (wiślański)
charakter grodu w Naszacowicach, czy wcześniejsze nieco, bo już z VI wieku,
słowiańskie osadnictwo otwarte, odkryte nad potokiem Baryczka w Podegrodziu? Tam
archeolodzy znaleźli zabytki bardzo ubogiej i prymitywnej ceramiki tzw. kultury praskiej,
z tego powodu wiązanej
dotychczas przez archeologię tylko z wczesnymi Słowianami. Właśnie znaczna część rosyjskich, ukraińskich,
a zwłaszcza polskich archeologów (najpierw prof. K. Godłowski, a obecne zwłaszcza M. Parczewski
i jego uczniowie z tzw. krakowskiej szkoły archeologicznej, allochtonicznej) przekonująco
udowadniała, że praojczyzną Słowian były bagniste ziemie na Ukrainie - w
dorzeczach środkowego Dniepru, Prypeci i górnego Bugu, gdzie przez wieki żyli w odcięciu od świata
i jego wytworów, i dopiero w V/VI wieku wyruszyli w poszukiwaniu lepszych siedzib
i tak ze swoją prymitywna kulturą weszli na arenę dziejową.
O Słowianach
więc, zgodnie z jakąś dziwną "poprawnością polityczną",
do niedawna mówiono, że:
"Oni
[Słowianie] bardzo późno, na tle Germanów, zeszli z drzew w puszczy nad Dnieprem"
i zajęli ziemie nad Wisłą, od niepamiętnych czasów germańskie.
albo: "Słowianki zbyt późno wyłoniły się z bagien nad
Prypecią!",
czy też: "Tylko Cyganie później od Słowian pojawili się na
arenie dziejowej".
Czy na pewno tak było? Czy taka jest cała prawda?
To pytanie legło u podstaw powstania tego portalu, poświęconego
genealogii najpierw dokumentalnej niektórych moich lub
parafialnych rodów, a następnie
genetycznej niektórych osób i grup, jak Polaków, Słowian,
Europejczyków i Praindoeuropejczyków
oraz Y-chromosomalnego Adama, jako ojca całej współcześnie
żyjącej ludzkości. W międzyczasie genealogia genetyczna
poczyniła milowe postępy i, niestety, niektóre pierwsze
konstatacje stały się przestarzałe lub wprost nieużyteczne,
oczekują więc na poprawę lub uzupełnienia (10.05.2016) .
1.
"GENEalogia Y-DNA prawdę ci powie!" o początku
i pokrewieństwie ludów na kuli ziemskiej
(haplogrupy
Y-DNA)
Oto pojawia się genetyka, a właściwie nie
genetyka, lecz genealogia Y-DNA, ze swoimi
fascynującymi możliwościami dla badaczy tych zagadnień! M.in. dostarcza ona
danych, które umożliwiają umiejscowienie konkretnego człowieka na drzewie
genealogicznym rodu, plemienia, wielkiego ludu lub nawet całej ludzkości.
To
dlatego, że w męskim chromosomie Y przechowuje się i zasadniczo wiernie dziedziczy w
trakcie przekazywania życia z ojca na syna materiał
DNA wraz z jego historycznymi zmianami, zwanymi mutacjami.
Męski charakter genealogii Y-DNA jest istotny w ustalaniu pierwotnej grupy
rodowej, plemiennej, czyli pierwotnego etnosu (gr. "etnos" to rój, trzoda,
plemię). Bowiem w cywilizacji patriarchalnej ojcowski rodowód był istotnym czynnikiem
genezy i rozwoju grup rodowych, plemiennych i etnicznych. Płciowy chromosom Y,
choć nie służy przekazywaniu cech somatycznych (z wyjątkiem płciowych) ani tzw. rasowych, zawiera w
sobie jakby bibliotekę rodowodu ojcowskiego wszystkich poprzednich pokoleń w
linii prostej. W genealogii Y-DNA
bierze się pod uwagę te mutacje, które dokonują się w niekodujących (niegenowych)
odcinkach Y-DNA
(ale to około 95% całej nici DNA!).
Mianowicie
spośród około 57 milionów możliwych w chromosomie Y mutacji, niektóre,
np. SNP (Single Nucleotide Polimophism; snip) dokonują się
bardzo rzadko, bo w poszczególnych parach zasad (nukleotydów)
przeciętnie raz na miliard lat, a więc praktycznie najwyżej raz w dziejach
ludzkości i
tylko u jednego mężczyzny, spontanicznie, jakby losowo, jako błąd w
procesie kopiowania Y-DNA. Są one zasadniczo dokładnie przekazywane
w trakcie zapłodnienia (mejozy) wszystkim męskim potomkom. Przy tym
jednak, choć rzadko, może dojść do kolejnej mutacji, choć w
innym miejscu (markerze), która zostaje przekazana potomstwu
podobnie jak i poprzednie.
Mutacje
SNP, przekazane męskim
potomkom, są przez genetykę oznaczane odpowiednią literą, wskazującą na
laboratorium, gdzie mutacje odkryto i cyfrą, oznaczającą
kolejną liczbę odkrycia mutacji w tym laboratorium. Stąd np.
takie symbole mutacji SNP: M417, L260, P45, CTS1211, DF93 itd.
 |
Komputerowy obraz mutacji typu SNP (single nucleotide
polomorphism
Symbole zasad/nukleotydów: A
adenina, C cytozyna, G guanina,
T tymina.
Widzimy dwie sekwencje (dwa uklady) par zasad:
wyżej, w próbce odniesienia jest sekwencja: GATTGGATGGGGT
na dole, w próbce badanej sekwencja nieco inna: GATTGGTTGGGGT
bowiem pojawiła się zasada T (tymina) tam,
gdzie w próbce odniesienia (w linii na górze)
jest A (adenina), czyli Adenina
została zastąpiona Tyminą.
Oznaczenie mutacji:
A>T |
Od mutacji SNP zaczyna istnienie nowy
ród (lub podród) genetyczny ojcowski (męski), a od mutacji mtDNA
- genetyczny ród matczyny (żeński), zwany haplogrupą (h.g.) Y-DNA człowieka
lub mtDNA (gr. 'haplous' jednolity, pojedynczy).
Haplogrupa więc to grupa ludzka, pochodząca od jednego
mężczyzny lub kobiety, u których w trakcie poczęcia doszło do określonej
mutacji zwanej SNP; mutacja ta występuje niezmiennie (gr. haplous) u wszystkich jego męskich
potomków, nawet mimo pojawienia się u nich kolejnych mutacji
(podobnie jak mutacje mtDNA u potomków żeńskich).
Haplogrupy oznaczamy umownie kolejnymi literami alfabetu łacińskiego,
np. męskie - od A do T, oraz dodatkowo
cyframi 1, 2, 3... oraz literami a, b, c... , jak np. R1a1a1 (jak aktualnie
prasłowiańska i praindoirańska) albo R1b1a2 (jak podstawowa
grupa genowa w Europie Zachodniej, zwana tez celtycką lub
atlantycką).
Natomiast pierwszy mężczyzna, przodek wszystkich obecnie żyjących mężczyzn na
świecie, tzw. Y-Adam, został oznaczony literą A. Potwierdziła się
bowiem zapisana w Biblii tradycja o jednym człowieku u początku ludzkości,
zwanym w genealogii genetycznej Y-chromosomalnym Adamem lub
Y-Adamem. Pierwszego potwierdzenia
dokonały badania
Cruciani et al., maj 2011,
który wykrył szereg mutacji SNP oznaczone później jako
haplogrupa A1, dziedziczone głównie przez wszystkie haplogrupy
pozaafrykańskie; oraz mutacje oznaczone jako bratnia haplogrupa
A0, wykryte w Kamerunie i dziedziczone tylko przez tę grupę
afrykańską. Z kolei w roku 2012 z
inicjatywy grupy klientów komercyjnej firmy FTDNA (tzw. "citizen
sciencists") i na ich koszt dokonano kolejnej serii badań,
które doprowadziły do wykrycia kolejnych osób z grupy A0, co
umożliwiło zatwierdzenie wyników Crucianiego, a nadto daleko
starszej haplogrupy, oznakowanej jako A00. Dziedziczna jest
głównie przez nieliczną ludność żyjąca w południowo-zachodnim
Kamerunie, wśród plemion Bangwa i Nkongho-Mbo. Paralelna
do A00, bratnia grupa mutacji SNP, otrzymała oznaczenie A0T,
gdyż jest dziedziczona przez wszystkie haplogrupy afrykańskie: A
(oprócz A00) i B oraz pozaafrykańskie od C do litery T. Liczba
SNP umożliwia datowanie tych haplogrup, a więc i ich wspólnego
przodka, Y-Adama, na około 200.000 lat (w pracy
opublikowanej przez Mendez et al. 2013 Y-Adama datowano nawet na
około 338.000 lat!).
A oto laboratoryjna "metryka" jednej z
tysięcy mutacji (SNP) Y-Adama, przekazanych obydwu liniom
ludzkości - A00 i A0T - jako wynik zmiany zasady
cytozyny w tyminę, czyli C->T. Odkryta jako pierwsza z tej
grupy stała się symbolem Y-Adama (w 2012-2013 r. liczba znanych SNP
Y-Adama wynosi już 1109):
Powyższe badania umożliwiły opracowanie nowego diagramu
filogenetycznego drzewa współczesnego człowieka.
|
Drzewo genealogiczne
współczesnej ludzkości.
(otwórz w
pdf)
Stan wiedzy:
10 listopada 2012
i 2013 r.
Porównaj:
Nowe drzewo genealogiczne
mtDNA |
Autor i kilku jego
bliskich aktywnie uczestniczyli w inicjatywie, programowaniu i ponoszeniu
kosztów testowania najstarszych, do niedawna nieznanych albo
uważanych za odrębne od reszty ludzkości, rodów afrykańskich,
celem ustalenia istnienia, czasu i miejsca chromosomalnego Adama, ojca
całej dzisiaj żyjącej ludzkości.
10.11.2012 ogłoszono w USA wyniki badań
klanu Perry'ego, rodem z Fontem w zachodnim Kamerunie.
Jest to gałąź
współczesnego człowieka, która najwcześniej wyłoniła się w
rodzie Y-Adama. Dzięki temu przodka całej ludzkości, Y-Adama, datuje
się obecnie albo na 338.000 (Hammer 2012; Mendez et al. 2013) lat, licząc
jednak po 30 lat na jedno pokolenie
(według dzisiejszego stanu). |
|
Po
koniecznej
poprawce,
na podstawie drzewa Scozzari et al. 2014, czas Y-Adama
wynosi około 320.000 lat.
Inne badania ukazują czas około
250.000 lat |
Aktualne drzewo genealogiczne genetycznych
rodów ojcowskich
współczesnych ludzi. Wrzesień 2015 r.

Części drzewa genealogicznego: afrykańska i
pozaafrykańska.
"Out of Africa" - około 68.500 lat temu

By user:Maulucioni. This work is derived from File:Yap tree.gif
by user:Wapondaponda - wikipedia, CC BY-SA 3.0,
https://commons.wikimedia.org/w/index.php?curid=1047118
- - -
Zobacz Tabelę czasu haplogrup i
populacji świata
http://www.tropie.tarnow.opoka.org.pl/hg_czas.pdf
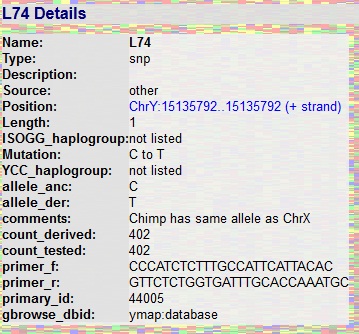
Zobacz procentowe rozmieszczenie poszczególnych haplogrup na
mapie świata.
http://www.scs.illinois.edu/~mcdonald/WorldHaplogroupsMaps.pdf
oraz: mapy rozmieszczenia w
świecie poszczególnych haplogrup
Por. datowanie węzłów
międzygrupowych (Marco Heinla, 2011):
http://beforepresent.dyndns.info/timevalues.html
oraz sporządzone przez Dienekesa Pontikosa i opublikowane na
jego blogu:
http://dienekes.blogspot.com/2012/08/dates-of-major-clades-of-y-chromosome.html
...............................................
Potwierdziła
się także tradycja o wspólnej matce wszystkich żyjących na świecie kobiet. Bowiem
dzięki mechanizmowi wiernego przekazywania potomstwu tylko przez matkę genu mitochoindrialnego, mt-DNA, regulującego powstanie i
działanie mitochondriów komórkowych, służących "energetyce
organizmu" oraz
dzięki mutacjom, które w trakcie kopiowania w tym genie zachodzą, można
dojść do pierwszej matki w linii prostej, czyli tzw. mitochondrialnej Ewy.
W genealogii genetycznej datuje się ją obecnie na około
177 tysięcy lat temu
(Behar et al. 2012) i też w środkowo-wschodniej Afryce.
Zespół
mutacji mitochondrialnej Ewy oznaczono symbolem
RSRS
- skrót od Reconstructed Sapiens Reference Sequence.
W ten sposób, korzystając ze
znajomości drzewa genealogicznego mutacji w Y-DNA i w mtDNA,
można genealogię swojego rodu, znaną np. z dokumentów i tradycji
rodzinnej, uzupełnić genealogią genetyczną - aż do pierwszych
praojców: chromosomalnego "Adama" i mitochondrialnej "Ewy".
 |
Ilustracja dziedziczenia
genów
mitochondrialnych (mtDNA) tylko w prostej
linii matczynej
i Y-DNA
tylko w prostej linii ojcowskiej.
Pozostałe geny,
tzw. autosomalne, dziedziczone są
tak w linii prostej,
jak i w liniach bocznych:
do ojców za pośrednictwem ich matek, a do matek za
pośrednictwem ich ojców.
dna-forums |
http://www.nottingham.ac.uk/-sczsteve/Gothenburg_13Oct2011.pdf
Zob. przykład takiej genealogii
ojcowskiej
Y-DNA
i matczynej
mtDNA
pewnej
rodziny.
Ze względu na to, że w naszej
cywilizacji patriarchalnej (ojcowskiej, męskiej) pierwotne grupy etniczne mają genezę patriarchalną,
co potwierdza archeogenetyka, interesuje nas szczególnie ustalenie pokrewieństwa
i pochodzenia wspólnot plemiennych i etnicznych przez śledzenie dziedzictwa genetycznego w
męskim chromosomie Y-DNA. Zajmuje się tym genealogia Y-NA,
zwana też, choć trochę niewłaściwie, genogenealogią. Genealogia Y-DNA bywa w
antropologii niekiedy mylona z pokrewną nauką, zwaną genetyką
populacyjną, której zadaniem jest śledzić genetyczne
podobieństwa między dzisiejszymi społecznościami, z całą ich
biologiczno-genową złożonością.
Odkrycia i datowanie "Człowieka z
Ust'-Ishim" u uścia' (sic!) rzeki Iszym
do Irtyszu w zachodniej Syberii, a także haplogrupy K2*
(Fu et al., 2014)
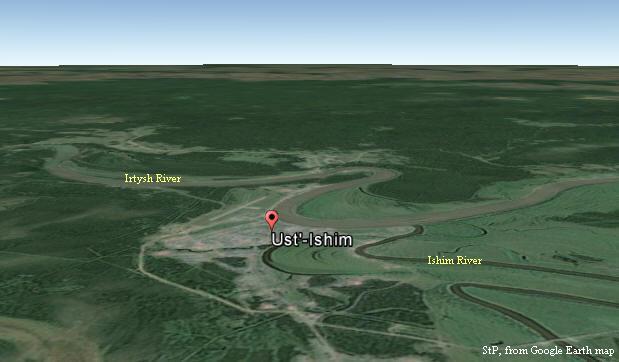
Tam, w Ust'-Ishim nad rzeką Irtysz, 45.000 lat temu żył
"Czlowiek z Ust'-Ishim", potomek haplogrupy K2*,
żyjącej około 50.000 lat temu, która wcześniej migrowała w
kierunku Sundalandu, gdzie wnet pojawił się inny jej potomek,
haplogrupa MP, praprzodek większości Europejczyków (zob. tabela
Czasu rozwoju haplogrup Y-DNA).
Tempo mutacji (SNP) w Y-DNA 0,76x10^-9
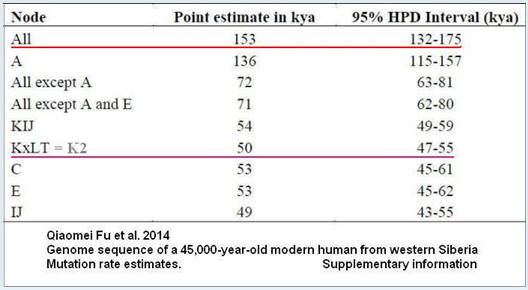
Uwaga.
Zaniżone datowanie wszystkich (All) haplogrup człowieka
współczesnego (153 tysiące lat zamiast około 275 lub
przynajmniej 200 tysięcy, jak do niedawna datowano) jest
wynikiem braku pełnego testowania ludzi haplogrupy A00.
2.
Genealogia Y-DNA POWIE CI też o bliższym pokrewieństwie
rodów, rodzin i osób
(Haplotypy
Y-DNA)
Oprócz omówionych wyżej mutacji SNP, w męskim Y-DNA
dokonują się jeszcze inne, mniej czytelne w genealogii, ale
daleko częstsze zmiany. Polegają one na zmiennej ilości
powtórzeń odpowiedniego tandemu (sekwencji) zasad, np. ATGC,
w pozagenowych mikrosatelitarnych markerach Y-DNA,
nazywanych STR (Short
Tandem Repeats, czyli powtarzenia krótkiego tandemu, czyli
sekwencji, zasad A-T-G-C). Ulegające zmianom miejsca, w których
dokonują się owe powtórzenia, nazywane są "loci" lub "markery" i oznaczane
są skrótem Dys# i numerem tego miejsca; zaś zmieniona liczba powtórzeń owych
tandemów stanowi liczbę alleli w danym markerze (miejscu),
np. DYS393=13. Zestaw liczb
owych powtórzeń (alleli) u jednego mężczyzny stanowi jego
haplotyp (ht) STR.
Haplotyp, czyli zestaw ilości mutacji w poszczególnych miejscach (markerach) Y-STR
jednego mężczyzny, umożliwia porównanie z haplotypem innego mężczyzny z tej
samej haplogrupy,
przeliczenie dzielących ich mutacji czyli GD (genetyczny dystans), ustalenie na
ich podstawie wspólnego najbliższego przodka (MRCA) i rozpoznawanie bliskości
ich pokrewieństwa. Bywa to
wykorzystywane np. w sądownictwie (dla ustalenia lub wykluczenia ojcostwa) albo
w budowie drzewa genealogicznego jakiegoś wielkiego rodu.
Spotykane liczby alleli w
poszczególnych 67 markerach (loci), w standardowej kolejności
firmy FTDNA i w najczęstszych haplogrupach w Europie
przedstawione są na na specjalnej
tablicy, a zwłaszcza na naszej stronie:
Dyskusja o mutacjach.
Od 5 kwietnia 2011 r. jako
standard uważa się testowanie nie 67, a 111 markerów w firmie
FamilyTree DNA.
Zestaw podobnych haplotypów STR
może być niekiedy użyty bez bardziej kosztownych badań, choć z
daleko mniejszą pewnością, do ustalenia haplogrupy SNP, w ramach której
owe haplotypy powstały. Jednak trzeba wiedzieć, że podobne haplotypy mogą pojawiać
się równolegle w rozmaitych haplogrupach.
Oto przykład 25-markerowego haplotypu modalnego, czyli najczęstszego, słowiańskiej haplogrupy
R1a1a w krajach Europy Środkowej i Wschodniej:
zwłaszcza: Polska, Ukraina, Rosja, Białoruś, Litwa, Niemcy,
Czechy, Słowacja i Węgry (według Wikipedii)
|
DYS |
393 |
390 |
19 |
391 |
385A |
385B |
426 |
388 |
439 |
389I |
392 |
389II |
458 |
459A |
459B |
455 |
454 |
447 |
437 |
448 |
449 |
464A |
464B |
464C |
464D |
|
Allele |
13 |
25 |
16 |
10 |
11 |
14 |
12 |
12 |
11 |
13 |
11 |
30 |
16 |
9 |
10 |
11 |
11 |
23 |
14 |
20 |
32 |
12 |
15 |
15 |
16 |
Liczba DYS w programach FTDNA oznacza numerowane
miejsce (marker) w STR męskiego Y-DNA.
Allele wyrażają ilość powtórzeń określonej sekwencji
(tandemu, następstwa) zasad A, C, G, T w danym miejscu Y-DNA.
Np. na podstawie Dys 385a=11, 439=11, 447=23 rozpoznaje się powyższego haplotypu
haplogrupę R1a1a1g, typ N
(kraje słowiańskie).
Powstanie w drodze mutacji
(np. przestawienia kolejności zasad A,C,G,T) nowych haplotypów w
ramach jednej haplogrupy wymaga czasu. Stąd ilość tych
mutacji w Y-STR bywa wykorzystywana w ramach genealogii Y-DNA do obliczania czasu od wspólnego przodka, w
skrócie TMRCA (Time to the Most Recent Common Ancestor). Metodami statystycznymi ustalono, że:
1) w przypadku przeliczenia alleli w 12-markerowym
haplotypie należy się spodziewać
0,022 mutacji na 25 lat, co daje jedną pełną mutację na haplotyp na 1136 lat.
2) w przypadku przeliczenia alleli w 25-markerowym
haplotypie należy się spodziewać 0.046
mutacji na 25 lat, co daje jedną pełną mutację na haplotyp na 543 lata.
3) w przypadku przeliczenia alleli w 37-markerowym
haplotypie należy spodziewać się 0.09
mutacji na 25 lat, co daje jedną pełną mutację na haplotyp na około
278 lat.
4) w przypadku przeliczenia alleli w 67-markerowym
haplotypie należy spodziewać się 0,12 mutacji na
25 lat, co daje jedną pełną mutację na haplotyp na około
208 lat (stawki Klosowa-Rożanskiego w: Wiestnik 3/12, 2010 r.).
|
UWAGA!
Po sprawdzeniu wielu
zagranicznych publikacji naukowych i śledząc naukowe
fora poświęcone genealogii Y-DNA, zwłaszcza RootsWeb,
GeneDiversitas, MolGen lub Rodstvo, dochodzę do
przekonania o dość znacznej wiarygodności rosyjskiego
środowiska naukowego, któremu przewodzi
rosyjsko-amerykański naukowiec, prof. Harwardu
Anatol KLOSOW
(ang. Anatole Klyosov, ros. Анатолий A. Клёсов,
tu w skrócie: A.K.; Moskwa-Boston-Massachussetts), biochemik,
specjalista od kinetyki chemicznej, autor
uniwersyteckiego podręcznika w tej dziedzinie oraz
bardzo licznych i całościowych prac z dziedziny
genealogii Y-DNA. Obecnie z powodzeniem udoskonala i
propaguje on naukowe zasady liczenia mutacji STR, budowy
genealogicznego drzewa Y-DNA i obliczania czasu do
wspólnego przodka populacji danej haplogrupy (TMRCA).
Ostatnio w analizie haplotypów
i ostrożnym datowaniu populacji wyróżnia się też naukowiec-biochemik japoński
ukraińskiego pochodzenia, Igor Rożanski, cieszący się
środkowoeuropejską haplogrupą R1a1a1g-M458. Jego dziełem jest
zestaw cennych analiz i map drzewa genealogicznego R1a1a1, z
których korzysta się na tej stronie. |
|
Wykładniczy czas mutacji
widocznych (obserwowalnych)
Dotąd bardzo
trudny problem rozpoznawania ilości mutacji powtórnych /
zwrotnych, pojawiających się w markerach szybciej mutujących,
zupełnie niewidocznych i niedających się odczytać, który zmuszał autorów do
wykonywania rozmaitej akrobatyki matematycznej, stara się
rozwiązać najnowsza tabela Pełnego Czasu Mutacji (
PCM / FTM, zob.), utworzona na naszych stronach
i opublikowana na niektórych międzynarodowych forach naukowej genealogii
genetycznej.
Przedstawia ona nie liniowy, a
wykładniczy ciąg czasu mutacji, starając się uwzględnić
współczynniki mutacji niewidocznych, narastających wykładniczo
wraz z każdą mutacją widoczną. Na skutek tego, podczas gdy
pierwszej mutacji widocznej przypisuje się statystycznie 111
lat, to sześćdziesiątej już 22 tysiące lat. Liczenie czasu
kolejnych mutacji widocznych staje się już praktycznie
niemożliwe.
Szerszy opis problemów,
związanych z obliczaniem liczby mutacji i ich czasu, zob. na
stronie Dyskusja o mutacjach. |
Obecnie każdy człowiek, przekazując wymaz (z jamy ustnej) ze swoim DNA, może wziąć udział, z
niewielkim wydatkiem pieniężnym, w
programie tych badań (program FamilyTreeDNA). Zob.
Polish project, który administruje Amerykanin polskiego
pochodzenia, L. Mayka. Krótki poradnik zob. niżej, w
rozdziale 16.
 |
Dolna część drzewa genealogicznego ludów
Europy, tylko haplogrupy męskie Y-DNA.
Od lewej - haplogrupy dzisiejszych ludów:
R1b, R1a1, Q1a, N1c1, I1a, I2a2, J2b, J1, G2a3, E-V,
E-M.
O nich mowa niżej. W centrum - czas powstania
(w latach) odpowiednich mutacji Y-SNP czyli początku poszczególnych haplogrup. (Eupedia-tree2) |
3.
jak powstały ludy pozaafrykańskie?
Mapa ogólna rozprzestrzeniania się współczesnego człowieka z
Afryki do Eurazji i dalej.
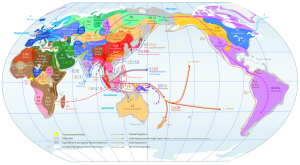
Migracje i rozmieszczenie haplogrup Y-DNA w świecie (kliknij,
powiększ!)
Zob. miejsce haplogrupy A00 i A0 w Kamerunie; tam początek
współczesnego człowieka.
źródło:
https://en.wikipedia.org/wiki/File:World_Map_of_Y-DNA_Haplogroups.png
Zob. pochodzenie rdzennych Amerykanów,Ri Q:
http://www.sciencemag.org/content/342/6157/409.full
(Zobacz też:
Pochodzenie ludności w rejonie
Dunajca)
https://dnaexplained.files.wordpress.com/2014/10/hammer-2014-30.jpg?w=584&h=380
Pierwsze
słynne "Out of Africa"
współczesnego człowieka na Półwysep Arabski mogło dokonać się
wnet po 100.000 lat temu, o czym mają świadczyć niektóre
świadectwa archeologiczne. W tym okresie, zapewne w Afryce,
powstaje haplogrupa BT*, której
liczne SNP są dziedziczone przez wszystkie pozaafrykańskie
haplogrupy. Na czas około 70.000 lat temu datuje
się wielką katastrofę przyrodniczą, związaną z wybuchem wulkanu
Toba w Indonezji. Musiała ona dotknąć i najstarsze populacje
ludzkie, migrujące w kierunku Azji. Widoczne jest to także na haplogrupie BT, której liczne mutacje w jednej linii ojcowskiej
świadczą o przeżytym wąskim gardle (bottleneck) demografii.
Przeżył tylko jeden przodek, oznaczony dziś jako haplogrupa BT,
który otrzymał szansę być ojcem prawie całej pozaafrykańskiej
ludności świata.
Zapewne poza Afryką, około
70.000 lat temu, już poza Afryką
haplogrupa
CT rozgałęziła się na DE i CF.
Hg E - po wydzieleniu siĘ wnet
powróciła do Afryki, gdzie się bardzo upowszechniła i stanowi
większość ludów tego kontynentu. Hg
D - migrowała daleko do Azji
środkowej i wschodniej (Japonia). Potomkowie z hg
C żyją w krajach Azji zachodniej,
południowej i południowo-wschodniej, nadto jako aborygeni w
Australii oraz pierwotna ludność na wyspach Polinezji i
nielicznie w Ameryce.
Natomiast potomstwo naszego przodka z haplogrupą F dało początek prawie 90% pozaafrykańskiej ludności
świata. W gronie potomków haplogrupy F
(naszego przodka,
zob.)
w Azji Mniejszej wyłoniła się najpierw współna haplogrupa GHIJK,
a wnet potem żyjąca dziś głównie na
Zakaukaziu i nielicznie w
Europie haplogrupa G (wyróżniająca dziś głównie Ormian).
Haplogrupa G na początku VI tysiąclecia przed Chr. przyniosła do
Europy rolnictwo. Natomiast z synowskiej haplogrupy HIJK
wyłoniła się haplogrupa H. To żyjący dziś w Indiach, a potem
ich część także w Europie jako Cyganie/Romowie.
Od około 55
tysięcy lat temu Azji Mniejszej z ojcowskiej grupy IJK rozwija się populacja z haplogrupą IJ,
a z niej
potem wyłoni się w Europie ludność staroeuropejska, "przedindoeuropejska", z haplogrupą I,
którą by trzeba utożsamić prawdopodobnie ze znanymi w Europie populacjami
przeważnie pod
nazwą Wenetowie/Wenedowie. W południowej Skandynawii dali oni
początek etnosowi pragermańskiemu. Natomiast w Azji Mniejszej
powstała haplogrupa J, głównie semicka.
W czerwcu
roku 2014 T. Karafet et al. ogłosili wyniki badań ważnych dla
ustalenia, dotąd raczej enigmatycznie nakreślanego, drzewa
filogenetycznego w pionie haplogrup A-R, w jego młodszych
gałęziach:
Improved
phylogenetic resolution and rapid diversification of
Y-chromosome haplogroup K-M526 in Southeast Asia,
(Link i dostęp w:
http://dienekes.blogspot.com/2014/06/refined-structure-in-haplogroup-k-m526.html
).
...............................
Nota z dnia 25.08.2021.
Ważnym przyczynkiem w temacie pochodzenia wczenych gałęzi drzewa
współczesnego człowieka jest praca Hallast
P. et al. 2021, Pochodzenie z Azji Południowo-Wschodniej dla
współczesnych nieafrykańskich ludzkich chromosomów Y (z ang.)
(
https://link.springer.com/article/10.1007/s00439-020-02204-9
)
Autorzy przekonująco wykazują, że po prawdopodobnym podziale w
haplogrupie CT jeszcze przed wyjściem z Afryki, wnet po roku
54.000 lat temu dokonała się szybka ekspansja w kierunku
południowo-wschodniej Azji trzech haplogrup, C, D i FT.
Genetyczne ślady ich zgrupowania widzimy na poniższej mapie
(kolor czerwony) na terenie tzw. Sundalandu. Stamtąd dopiero, po
rozpadzie haplogrupy FT na F i GHIJK, ta ostatnia w swoich
podgrupach jeszcze przed rokiem 50.000 wydzieliła z siebie 15
linii, z których czternaście rozmieściło się wówczas w
południowo-wschodniej i południowej Azji lub Oceanii,
ekspandując potem na całą Eurazję i pozostały świat.
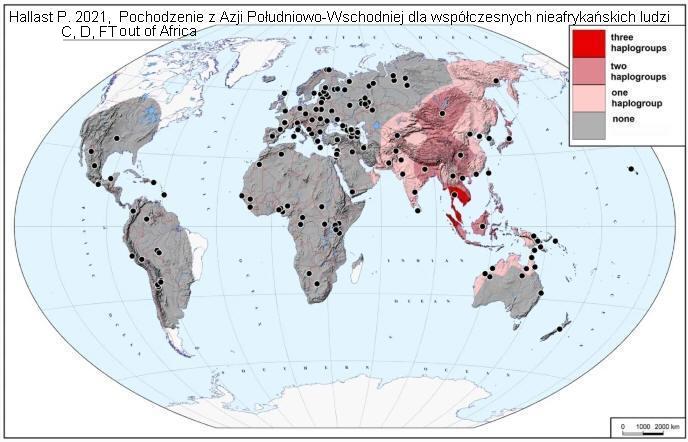
Hallast P. 2021, Pochodzenie z Azji
Południowo-Wschodniej dla współczesnych nieafrykańskich - post
Out of Africa
Ta migracja naszych praojców z
Afryki w trzech grupach: C, D i FT do Sundalandu i potem w
wydzielonych
tam i później grupach z Sundalandu ku Europie
jest ważnym epizodem w naszych dziejach rodowych.
0 - 0 - 0 - 0 - 0
Według Karafet 2014, w rodzie naszego praprzodka z haplogrupą K-M9, po wydzieleniu się
gdzieś w Azji Poludniowej podgrupy K1-P326, w której powstały
gałęzie L (żyjące dziś zwł. w Pakistanie, Indiach i Sri
Lance) i T (żyjąca dziś na Bliskim Wschodzie, pd.-zach. Afryce i w Europie),
w drodze ku Azji Południowo-Wschodniej wyłoniła się najpierw
mutacja K2-M526 (bliski przodek "człowieka a UST'-Ishim",
datowanego radiowęglowo na 45 tysięcy lat), a już zapewne na terenie
Sundalandu, (dziś Wyspy Sundajskie
w Indonezji) wówczas w całości
będącego na powierzchni, kolejna podgrupa z mutacją P331, oznaczana teraz jako
haplogrupa K2b, oznaczana także jako
MP.
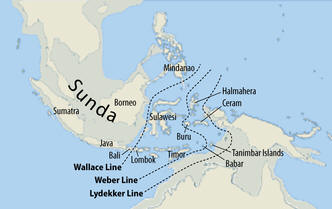
Sundaland i jego domniemane granice
południowe,
stosownie do zmiennego poziomu mórz (
http://en.wikipedia.org/wiki/Sundaland
)
Haplogrupa MP uległa tam szybkiemu rozwojowi demograficznemu. Zapewne pod
koniec okresu wielkiego zlodowacenia (LGM) około 18-15 tys. lat temu
i podniesienia się poziomu mórz zaszła konieczność ucieczki z Sundalandu
i emigracji euroazjatyckiej, w której brały udział powstające
haplogrupy Q i R.
Oto jak M. Hammer przedstawia
migrację od Sundalandu ku Europie:
https://dnaexplained.files.wordpress.com/2014/10/hammer-2014-30.jpg?w=584&h=380
Niektórzy potomkowie
haplogrupy Q poprzez północno-wschodnią
Syberię i Alaskę około 15.000 lat temu
zawędrowali na kontynent Amerykański, dając wraz z hg C początek
autochtonnym Indianom w Ameryce Północnej i Południowej.
Natomiast haplogrupa R-M207
powstała około 34.000 lat temu (zob. tabela) i rozproszyła się
gdzieś m.in. w rejonie Ałtaju. O tym świadczy jej potomek, tzw.
"Chłopiec z Malta (Mal'ta)", którego archeologiczne szczątki,
datowane na 24.000 lat temu, znaleziono we wsi Mal'ta, w
pobliżu Jeziora Bajkał. W tym rejonie mogło też dojść do wydzielenia się i migracji podgrup R1 i R2. Ta druga
zatrzymała się głównie na Półwyspie Indyjskim. Ta pierwsza, z
mutacją M173 i swoimi podgrupami R1a i
R1b, rozproszyła się głównie w Eurazji Środkowej i
Zachodniej, głównie w Europie.
Populacje haplogrupy R1 z jej podrupami
R1a i R1b, które zachowały pierwotny "ojcowski" język, są
twórcami i użytkownikami języków tzw. indoeuropejskich,
nazwanych tak ze względu na geograficzną ich rozległość od
Subkontynentu Indyjskiego na południu Azji po zachodnie krańce
kontynentu Europejskigo.
Uwaga.
Powyższe dane porównaj:
Chiaroni et al. 2009; tam też najnowsza mapa powstania poszczególnych haplogrup, ich
migracji oraz aktualnego rozmieszczenia ich populacji w świecie
(mapa).
Główne haplogrupy, R1a (kolor pomarańczowy) i R1b (kolor
czerwony) i inne w Europie
i na terenach przyległych na
południu i południowym wschodzie; według wikipedii, rok
2013.
|
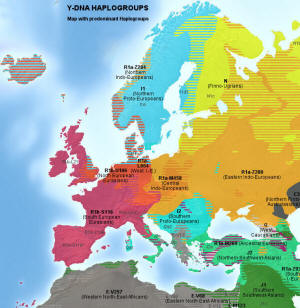
Główne haplogrupy Y-DNA w Europie
 (kliknij,
aby powiększyć) (kliknij,
aby powiększyć) |
Kolor żółty: haplogrupa N (N1b i
N1c) - grupy ugrofińskie
Pomarańcz.: haplogrupa R1a, grupa
rdzennie indoeuropejska
R1a-Z280 - wielka haplogrupa bałtosłowiańska
R1a-M458 - haplogrupa zachodniosłowiańska
R1a-L664 - haplogrupa Słowian Połabskich
R1a-Z284 - haplogrupa skandynawska
R1a-Z93 - haplogrupa południowo-wschodnia, azjatycka.
Niebieski: haplogrupa I (staroeuropejska)
11 - haplogrupa skandynawska, protogermańska
I2a2 - haplogrupa południowoeuropejska, dynarska
Czerwony: haplogrupa R1b
zachodnioeuropejska,
ibero-celtycka,
pierwotnie nieindoeuropejska
R1b-U106 haplogrupa anglo-saska
R1b-S116 (=P312) - haplogrupa celtycka
R1b- S116* - grupa iberyjska
R1b-A196 - grupa pirenejska
R1b-U152 - haplogrupa italoceltycka
R1b-L21 - haplogrupa północnoceltycka
R1b-M269 - grupa ojcowska dla europejskich R1b.
Zielony: haplogrupa J,
bliskowschodnia, nieindoeuropejska
J1 - haplogrupa semicka
J2 - haplogrupa bliskowschodnia i śródziemnomorska
Szary: haplogrupa E, afrykańska;
E-V13 bałkańska |
4.
http://www.eupedia.com/europe/Haplogroup_R1b_Y-DNA.shtml
ITALO-celtyckA i germańska HAPLOGRUPA R1b
w Europie.
Początek ludów "zachodnioeuropejskich"
Vincent Vizachero na
geograficznej mapie (http://vizachero.com/R1b1/R-Map.png)
zobrazował nam podział rodu R-M207 na R1-M173 i R2. Haplogrupę R2
znajdujemy głównie w Indiach. Natomiast z rodu R1 ewoluowały rody R1a i
R1b. Potomstwo rodu R1a-M420 zaludnia dziś głównie ziemie
słowiańskie. Natomiast w R1b-M343 autor uwidocznił podział na
trzy gałęzi: gałąź afrykańską (R-V88 i R-V69; głównie w
Kamerunie), gałąź europejską (potomstwo grupy R-M73) i
bliskowschodnią (R-M269).
 |
Tak genealog Vincent Vizachero (V.V.)
na Bliskim Wschodzie /Azji Środkowej ilustruje kolebkę
powstania rodu R, i jego
potomstwa: R2 (głównie Indie) oraz R1 (głównie Europa).
Ród R1b rozgałęzia się głównie na mutacje:
M269-Palestyna, V69-Afryka Subsaharyjska, M73 i jej
podgrupy - Europa.
Natomiast migrację rodu R1a i jego podgrup - V.V.
widzi powyżej mórz Kaspijskiego i Czarnego. |
A więc w ojcowskiej haplogrupie
R1-M173 wyłonił się,
prawdopodobnie w Azji Centralnej, przodek większości dzisiejszych zachodnich
Europejczyków, z haplogrupą R1b-M343. Jej dzieje
i poszczególne populacje wstępnie datował w obszernej publikacji Anatolij
A. Klosow w periodyku
'Wiadomości Rosyjskiej Akademii Genealogii Y-DNA':
1.
Gablogruppa R1b (czast' 1)
w:
Wiestnik Rossijskoj Akademii DNK-Genealogii t.3, nr 2, luty 2010
2.
Gablogruppa R1b (czast' 2) w:
Wiestnik, t.3, nr 3, marzec 2010: .
Wydzielenie się haplogrupy
R1b z ojcowskiej R1 dokonało się około 29.000 lat temu, zapewne
jeszcze w
Azji Poludniowo-Wschodniej.
Mapa migracji haplogrupy R1b
(M343) z Azji do Europy. Powstanie europejskich podgrup: L51,
L11 oraz następnych (wg. Eupedii):
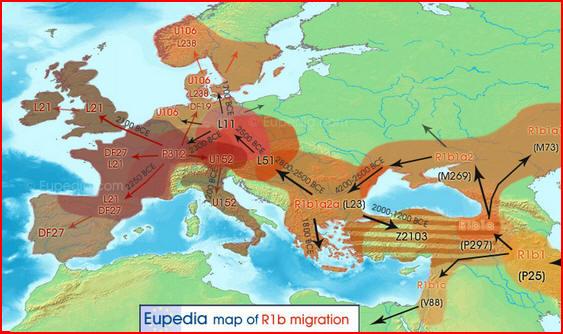
Procentowa i geograficzna częstotliwość
kluczowych zachodnioeuropejskich podgrup (mutacji) w haplogrupie R1b:
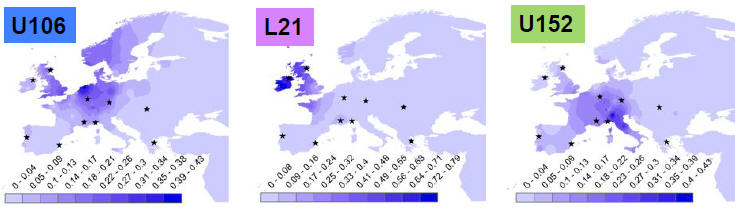
Euroazjatycka haplogrupa R1b(M343) i jej główne europejskie
podgrupy, pochodne europejskich L51 i L11.
1)
U-106 grupa germańska i anglo-saska. 2) L21 grupa celtycka.
3) U152
grupa italo-celtycka, alpejska.
(mapy z Eurogenes-Blog:
http://eurogenes.blogspot.com.au/2013_11_01_archive.html
Tam wskazanie źródło:
Michael Hammer, Origins of R-M269 Diversity
in Europe, University of Arizona, FamilyTreeDNA, 9th Annual
Conference).
Hammer 2013 powiedział: "Zespół dowodów potwierdza hipotezę, że rozkład
linii R-P311 jest wynikiem głównie
ruchów ludności, które wystąpiły dopiero po przybyciu rolnictwa w
neolicie. Populacje posiadające chromosomy R-P311 prawdopodobnie
zastały
Europę Zachodnią prawie opuszczoną przez neolityczne Y-DNA. To
spowodowało przewagę postneolitycznych przybyszów"
W Europie wspólny przodek (MRCA) podgrup haplogrupy
R1b pojawił się około 2500 lat a.C. (p.n.e.), co jest
najstarszym w Eurazji świadectwem jej istnienia. Jest to czas
kultury kurhanowej w Europie Środkowej i Wschodniej.
Warto wspomnieć, że osobna
gałaź haplogrupy R1b pojawia
się w Afryce Subsaharyjskiej, w okolicach
Czadu i Kamerunu (gdzie - wg pracy
F.Cruciani et al., 2010. - dochodzi do mutacji R-V88 i
jej podgrup) oraz Algierii, gdzie ich MRCA datuje się na około 1875
przed Chr.
Nieco odmienne, ale także godne zalecenia, są dane, które
prezentuje na swoich stronach
EUPEDIA (opis
haplogrupy i podgrup, datowanie, mapy, dyskusja). Według niej około 3000 lat przed Chr.
ludność hg R1b1a2, po całkowitym opuszczeniu kultury Majkop i częściowym
opuszczeniu Anatolii (zob.
mapa niżej), przeprawiła się
w okolice delty Dunaju, a potem dalej na zachód od Morza Czarnego. Tam zapewne
doszło do bliższego zetknięcia się z populacjami starszych haplogrup R1a1
(słowiańską), I1 i I2 (wenecką), J (bliskowschodnią) oraz E3
(afrykańską).
Później hg R1b1a2 przesunęła
się dalej wzdłuż Dunaju do jego źródeł, zatrzymując się na północ od Alp (dzisiejsza Austria i
południowe Niemcy): zobacz trasy migracji:
Eupedia.map
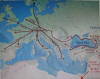 |
Migracja z Bliskiego Wschodu do
Europy zachodnioeuropejskiej
("celtyckiej") haplogrupy R1b
z Azji Mniejszej (Anatolia)
i Podkaukazia do rejonu pod
Alpami i dalej
na zachód, południe i północ.
Podano orientacyjne daty wydarzeń. Pełną mapę migracji zob.:
Eupedia |
Po kolejnych mutacjach w DNA, jak
wskazuje mapa, nastąpił także podział
"etniczny" dotychczasowej grupy celtyckiej na dwie zasadnicze podgrupy:
germańską (a raczej zgermanizowaną) R1b1a2a1a1-U106
i celtycką
R1b1a2a1a2-P312. Bowiem ta pierwsza
podgrupa,
pochodząca z ośrodków kultury halsztackiej, udając się ku północy i zajmując nadmorskie tereny
przy Morzu Północnym i przy zachodnim Bałtyku, po zlaniu się z grupą bałtycko/nordycką
(hg I1 i I2b), przeszła proces germanizacji.
Natomiast inne podgrupy celtyckie skierowały się
do Italii i na
półwysep Iberyjski (Baskowie, Iberowie - ok. 2250 p.n.e), do Galii (Francja)
2200, na Wyspy Brytyjskie (Irlandia 1800,
Brytania 400 p.n.e), Italia i Bałkany 1200, Nadrenia 1200, Skandynawia 1000
p.n.e. (datowanie wg Eupedii lub Klyosova)
Dziś cała zachodnia haplogrupa R1b1a2, ze wszystkimi swoimi
kilkunastoma mutacjami i podgrupami, stanowi w tych krajach podstawową populację, począwszy od
Skandynawii, gdzie regionalnie może być ok. 30% mieszkańców tej grupy, i Niemiec,
gdzie do całej grupy zachodnioeuropejskiej (tj. zgermanizowanych Celtów) należy blisko 40% społeczeństwa, aż
do Półwyspu Iberyjskiego (np. Baskowie) i
Wysp Brytyjskich,
które regionalnie nawet w około 80% wykazują się haplogrupą R1b1a2. Wysoki procent ludności z
tą haplogrupą żyje dziś także na kontynencie amerykańskim.
Długa migracja ludności
haplogrupy R1b1a2 z Azji Centralnej przez wysoko rozwinięte tereny ówczesnego
Bliskiego Wschodu, Azji Mniejszej i Półwyspu Bałkańskiego regionami południowymi
lub doliną Dunaju;
nadto kontakty z siostrzaną grupą
europejskich Słowian R1a1a1 i R1a1a1g, rozwijającą się wtedy już od
Krety i Bałkanów po Polskę i najbliższe ziemie stało się
olbrzymią szansą przyspieszonego rozwoju cywilizacji celtyckiej w Europie i stworzenia wcześniejszych, niż np. wśród Słowian, organizmów państwowych.
O udziale populacji celtyckiej
R1b w tworzeniu i rozwijaniu językowego i kulturowego
dziedzictwa indoeuropejskiego - zob.
w rozdziale 8.
O udziale Celtów w tworzeniu
archeologicznych kultur w starożytności - zob.
Dyskusja n.18.
Oto kolejna praca, Myres et al.
2010, na temat przybycia z Azji i zasiedlenia Europy przez
populacje haplogrupy R1b1a2 i jej podgrup w Europie:
|
Mapa przybycia do Europy
osadnictwa R1b1a2 i jego podgrup
 |
5.
http://www.eupedia.com/europe/Haplogroup_R1a_Y-DNA.shtml
Haplogrupa/RÓD
R1a
i jego główne podgrupy.
Ród zwłaszcza
Słowian i ich najbliższych krewnych, indo-irańskich Ariów.
Z genezą
Słowian jako etnosu (gr. "etnos" - plemię, ród)
sprawa jest niejasna, podobnie jak z innymi starożytnymi ludami.
Najstarszą słowiańską wersją ich nazwy zanotował Prokopius z
Cezarei: Spori, która skojarzyła sie mu ze słowem
sporadoi 'sporadyczni, rozproszeni'. Ale bliższa analiza
językoznawcza już wykazała, że słowiańskie słowo spori
oznaczało "sporzy, liczni".
(Odnośnie etnogenezy Słowian złą przysługę prawdzie naukowej
wyrządzili ostatnio skrajni allochtoniści, zwani kossinowcami-dnieprowcami,
którzy - na podstawie zacieśnionej definicji archeologicznej
kultury słowiańskiej - początek Słowian na ziemiach datują
jedynie na
VI w., ich praojczyznę lokują jedynie nad środkowym i górnym Dnieprem, a
ich genezę ustalają tam na czas niewiele wcześniejszy. Za nimi
poszli niektórzy polscy genetycy z kręgu medycyny sądowej, np.
K. Rębała i zesp. 2007, którzy badając pochodzenie dzisiejszych
Polaków, nie rozróżniają ich chromosomowych haplogrup SNP, a
biorąc pod uwagę tylko haplotypy STR, nieraz bardzo podobne mimo
odległych pochodzeniem haplogrup SNP, oceniają ich podobieństwo czy
różnorodność względem haplotypów innych populacji i na tej
podstawie orzekają o dacie ich genezy w... V czy VI w. i miejscu ich oddzielenia się od innych ludów - nad Dnieprem!
Podobna metoda, oparta na mieszaniu haplotypów rozmaitych
haplogrup i korzystaniu także z materiału autosomalnego,
zawiodła już wcześniej R. Płoskiego i zesp. 2002 do stwierdzenia, wbrew
naukowej genetyce, o wielkiej odrębności Polaków od innych ludów
słowiańskich; że pod "względem genetycznym jesteśmy prawie tak
samo oddaleni od Moskwy, jak i od Berlina").
W Rodzie R1a,
powstałym zapewne w zji Południowo-Wschodniej, na terenie dawnego
Sundalandu (zob. też niżej, tabela mutacji), w trakcie migracji ku Europie lub już w
Europie (Kujawy?) powstała kolejna mutacja SNP, oznaczona
symbolem M417 i określana obecnie jako haplogrupa
R1a1a1, uważana jest za ojcowską dla
indoeuropejskich
ludów; bowiem pod nią doszło do podziału tej rodziny genetyczne
na dwie wielkie grupy. Jedna, z mutacją Z93, dała początek indo-irańskim i azjatyckim Ariom i ich językowej odrębności.
Druga zaś, z mutacja Z283 dała początek wielu ludom europejskim,
a zwłaszcza Bałto-Słowianom i ich językom. Niektórzy naukowcy w
populacjach Z283 widzą genezę wszystkich indoeuropejskich
języków w Europie, także tych w europejskich haplogrupach R1b.

.
|
DRZEWO
mutacji Y-DNA w rodzie R1a. (Aktualizowano
01.08.2014)
Podstawa:
YFull experimantal tree,
Karafet 2014 oraz
aktualnych danych.
Datowwanie YBP oznacza rozwidlenie
h.grupy; Lata podano w przybliżeniu
R -
18.400 lat temu
M207/UTY2, P224, P227, P229, P232, P280, P285, S4, S8, S9,V45
•
R1
- 10.400 lat - M173/P241, M306/S1,P225, P231, P233, P234,
P236, P238, P242,P245, P286, P294.
...............................................................................................
• •
R1a - L62/M513,
L63/M511,
L145/M449,
L146/M420.
• • •
R1a1 -
L120/M516, L122/M448, M459,
Page65/SRY1532.2/SRY10831
• •
• •
R1a1a - M17, M198
.................................................................................................
• •
• • •
R1a1a1
- M417
-
7.700
lat temu.; mutacja i podział rodu indoeuropejskiego -
oddzielenie się CTS4385
• •
• • • •
R1a1a1a
-
CTS4385 i L664
- niewielka grupa europejska, północno-zachodnia
• •
• • • •
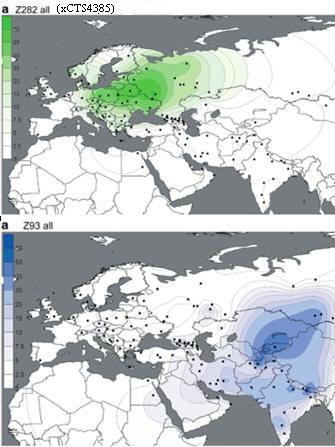
R1a1a1b - Z645; kolejny podział rodu indoeuropejskiego na Z283
(Europa) i Z93 (Azja)
................................................................................................
• •
• •
• • •
R1a1a1b1
-
Z283 -
powstanie rodu europejskiego.
• • •
• • •
• •
R1a1a1b1a -
Z282 - podział rodu europejskiego na gałęzi
M458, Z280 i Y2395.
................................................................................................
•
•
• • •
• • •
•
R1a1a1b1a1
- M458 -
Europa Środkowa. (i Zachodnia Ukraina),
• • • • •
• • • •
•
R1a1a1b1a1a
- L260
- gałąź zach.-słowiańska:
Polska, Czechy, Słowacja.
• • • • •
• • • •
•
R1a1a1b1a1b
- CTS11962
- gałąź zachodniosłowiańska.
• • • • • •
• • • •
•
R1a1a1b1a1b1
- L1029
................................................................................................
•
• •
• •
• • •
•
R1a1a1b1a2 - Z280
-
duża gałąź bałto-słowiańska;
•
•
• •
• • •
• • • R1a- Z92
- gałąź bałtyjska i wschodnia.
•
•
• •
• • •
• • • R1a- CTS1211
- gałąź "słowiańska"
• •
•
• •
• • •
• • • R1a-YP343
- środkowa Europa
• •
•
• •
• • •
• • • • R1a-P278.2
- gałąź "zachodnia karpacka".
• •
•
• •
• • •
• • • • R1a-YP380
- gałąź "stara karpacka".
• •
•
• •
• • •
• • • • • R1a-NN
• •
•
• •
• • •
• • • • • R1a-4700
• •
•
• •
• • •
• • • R1a-L784
• •
•
• •
• • •
• • • R1a-L1280
- północno-wschodnia europejska
• •
•
• •
• • •
• • • R1a-CTS3402 -
środkowa i wschodnia Europa,
gałąź bardzo
wsielka.
• • •
• •
• • •
• • • • R1a-Y237 -
• • •
• •
• • •
• • • • • R1a-YP419
• • •
• •
• • •
• • • • • R1a-NN
• • •
• •
• • •
• • • • • R1a-NN
• • •
• •
• • •
• • • • • R1a-YP234
• • •
• •
• • •
• • • • • • R1a-YP295
• • •
• •
• • •
• • • • • • • R1a-YP335
• • •
• •
• • •
• • • • • • • R1a-YPYP346, L366
- Pomorze
• • •
• •
• • •
• • • • • • R1a-L365
- Pomorze, Kaszubia
• • •
• •
• • •
• • • • • • • R1a-NN
• • •
• •
• • •
• • • • • • • R1a-NN
• • • • • • •
• •
• • • R1a-Y2613
• • • • • • •
• •
• • • R1a-Y33,
CTS8816
.................................................................................................
•
•
• •
• • •
• • R1a1a1b1a3
- Y2395 i Z284 -
gałęzie północno-zachodnioeuropejskie.
•
•
• • •
• •
• • • R1a1a1b1a3a
- S 5301 i L448 -
północna i
północno-zachodnia Europa
•
•
• • •
• •
• • • • R1a1a1b1a3a1
- CTS4179
• • • •
• • • •
• • • • R1a1a1b1a3a1a
- L176 -
Szkocja
• • •
• • • •
• • • R1a1a1b1a3b
- Z287 - starsza gałąź skandynawska
......................................................................................................
•
• • • •
• •
R1a1a1b2 - Z93 i Z94 -
powstanie gałęzi azjatyckiej i Aszkenazyjczyków
• •
• • •
• • •
•
R1a1a1b2a1 - L657
- zwłaszcza indo-irańscy Ariowie
• •
• • •
• • •
• • R1a1a1b2a1a
- Y7 - wśród Kazachów i Arabów
• •
• • •
• • •
• R1a1a1b2a1a2
- Z2124
- Eurazja (m.in. wśród Tatarów)
• •
• • •
• • •
• • R1a1a1b2a1a2a
- Z2123
• •
• • •
• • •
• • R1a1a1b2a1a2b
- Z2122 - głównie żydzi aszkenazyjscy,
lewici
• • •
• • •
• • •
• • R1a1a1b2a2b1
- F1345
• • • •
• • •
• • •
• • R1a1a1b2a2b1a
- CTS6 - aszkenazyjscy-lewici, z ok. 900 r. po Chr.
• • • •
• • •
• • •
• • R1a1a1b2a2b1b
- F2935
• • •
• • •
• • •
• • R1a1a1b2a2b2
- Y57
..........................................................
...............................................
...............................................
...........................................
• •
R1b
- M343, itd. - to początek wielkiego drzewa, bratniego
dla R1a; zwł. ludność środkowo i zachodnioeuropejska, zwłaszcza italo-celtycka
i germańska; w Europie od ok.
2500 lat
przed Chr.. Liczna też w Afryce Subsaharyjskiej (np. rejon Czadu
i pn. Kamerunu) V88.
.......................................................................................
.............................................
....................................................................
Uwagi
1. Obok symboli wypisano podstawowe mutacje SNP w Y-DNA,
określające haplogrupy.
2. Kolorem różowym oznaczyłem własne komentarze. Datowanie zasadniczo
w/g
http://www.familytreedna.com/public/R1a/default.aspx
3. Nazwy gałęzi zaczerpnięto z PolishProject, R1a1 and
Subclades i z drzewa YdnaIP autorstwa I. Rożanskiego, zob. niżej
4. Nie wszystkie firmy, jak np. światowa FTDNA, na
bieżąco aktualizują drzewo i wprowadzają
zmiany symboli,
stąd różnice i praktyczne
nieporozumienia.
|
Drzewo R1a
(z dnia 15.10.2013) - w uproszczeniu
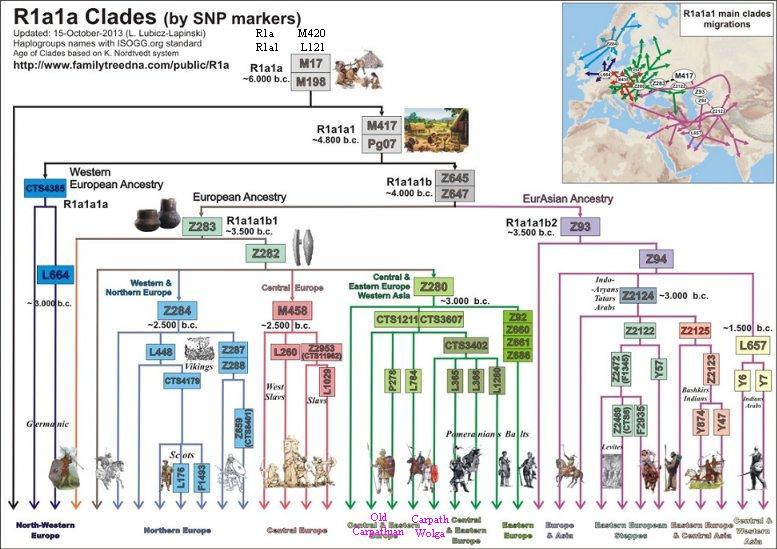
Opis powyższego drzewa
1) Haplogrupy R1a-L62 i R1b-M343
wydzieliły się spod R1 około 15.000
lat przed Chr., według nieostatecznego datowania.
2) Haplogrupa M17 (bez
mutacji M417) spotykana jest w ilości śladowej w pasie od Arabii
do Wysp Brytyjskich; czas ok.6000
lat przed Chr.
3) Haplogrupa M417 to
mutacja ojcowska dla małej grupy północno-zachodniej CTS4385 w
Europie i olbrzymiej Z645 euroazjatyckiej. Czas rozwidlenia
(split, bifurcation) megahaplogrupy M417 - około
4800 przed Chr., może w
Basenie Dunaju.
4) Haplogrupa Z645 około
4000 lat przed Chr. uległa
na skutek mutacji Z93 i Z283 oraz migracji ważnemu rozwidleniu
na grupę azjatycka i europejską. Rozejście się tych dwóch megagrup Z93 i Z283 z jednej wspólnoty praindoeuropejskiej
nastąpiło gdzieś w rejonie Morza Czarnego lub Morza
Kaspijskiego.
5) Haplogrupa Z93 dała
początek rodu azjatyckiego. To indo-irańscy Ariowie (Indie,
Pakistan, Afganistan Kurdystan), Tadżycy, Kirgizi i Żydzi
aszkenazyjscy (w Europie).
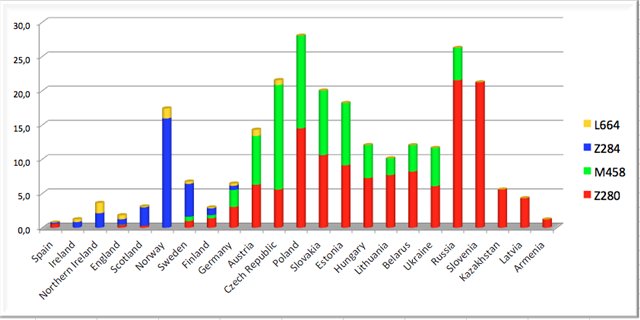
6) Haplogrupa Z283 to
ród europejski. Po kolejnej mutacji, Z282, około
3000 lat przed Chr. uległa
rozwidleniu na trzy ważne grupy z mutacjami: Z280, M458 i Z284.
6a) Haplogrupa Z280 jest
ogólnie zwana bałto-słowiańską. Jest najliczniejsza z powyższej
trójki. Jej obecność rozciąga się zasadniczo od Bałkanów do
Bałtyku i od Wołgi do Alp. Czas i przestrzenność
haplogrupy Z280 kojarzy sie z rozwojem archeologicznej kultury
ceramiki sznurowej (od 1290 lat przed Chr.). W składzie
haplogrupy Z280 znajdujemy m.in. grupy karpackie z mutacją
DYS452=28, pomorsko-kaszubską L365, wschodniopruską L366, bałtycką i
wschodniosłowiańską Z92, północno-wschodnią L1280 itd. .
6b) Haplogrupa M458, zwana
środkowoeuropejską; powstała prawdopodobnie na terenie
Czech (kultura unietycka?), ale rozciągnęła się przez Polskę do Ukrainy. W tej
grupie około 2500
lat przed Chr. powstała podgrupa L260 zwana
zachodniosłowiańską, a niekiedy nawet polską.
6c) Haplogrupa Z284, zwana
skandynawską, rozprzestrzeniała się głownie na terenie
południowej Skandynawii i zachodniej Norwegii (do 30 %
miejscowej ludności) oraz w niewielkim procencie na Wysp
Brytyjskich i Islandii. Ona zapewne wydajnie uczestniczyła w
indoeuropeizacji ludności germańskiej i celtyckiej.

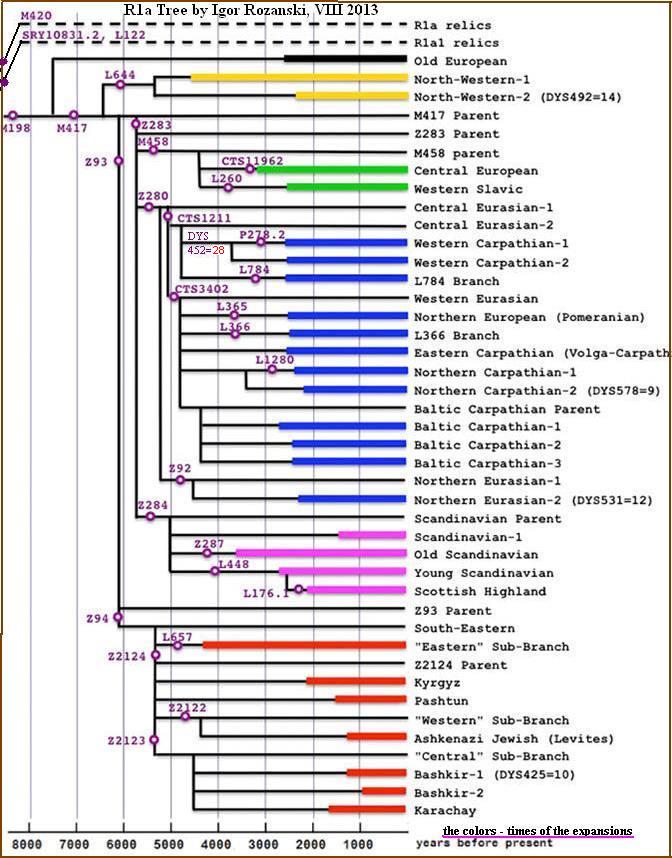
Drzewo rodu R1a od I. Rożanskiego
Sierpień 2013(
http://r1a.org/index.htm )
Główne mutacje i czas rozgałęzień i ekspansji. Nazwy przyjęte w środowisku Родство.
Ру
Prostopadłe linie wskazują czas rozgałęzienia. Kolory -
czas ekspansji (trwałego rozwoju demograficznego).
Ekspansja przestrzenna rodu R1a 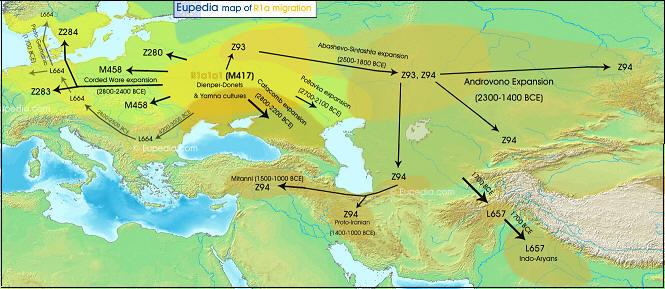
Ekspansja rodu R1a w Eurazji (według
portalu Eupedia)
Zadziwiająca jest
liczebność, a zarazem rozległość na mapie Eurazji
ludności z haplogrupą
R1a1a1-M417 z jej podgrupami): od Zachodniej Syberii i Kirgizji na wschodzie,
po linię Łaby i wschodnich Alp na zachodzie; od Bałtyku, a nawet wybrzeży
Norwegii i zachodniej Szkocji na północy, po Macedonię, Kurdystan, Iran,
Afganistan, Pakistan i Indie na południu!
Największą częstotliwość tej haplogrupy w Europie, do ponad 60% ludności, stwierdza się w pasie
między Bałtykiem a Morzem Czarnym, obejmującym całą Polskę i Zachodnią Ukrainę.
 |
Haplogrupa R1a1a1-M417
na mapie Eurazji
Widoczne zagęszczenia w rejonie Polski,
Ukrainy, Białorusi i Rosji, północnego Pakistanu i Indii oraz Azji
Środkowej (Kirgizja).
wg.
Chiaroni et al. 2009 |
Haplogrupą R1a1a1 wyróżniają się, jak widzimy, szeroko
rozprzestrzenione na kontynencie europejskim ludy słowiańskie z bałtyjskimi oraz te, które się dowodnie z ich wspólnoty
wydzieliły (łącznie z Kirgizami, Tadżykami i indoirańskimi Ariami (o czym niżej). Mając na uwadze to, że spośród
kulturowych wyróżników etnosu język jest
zasadniczo najbardziej trwałym dobrem, a według niezmiennej zasady, zakłócanej tylko
nadzwyczajnymi zdarzeniami,
język otrzymuje się w rodzinie od przodków i
praprzodków jako podstawowe dobro tradycji rodowej i plemiennej,
więc początek słowiańskiego języka wolno i trzeba łączyć z początkiem
nie tylko oddziału R-M458, ale i euroazjatyckiego R-M417, a nadto jego substratu trzeba dopatrywać się
już w ojcowskiej haplogrupie R1a (gdzieś
w Azji Środkowej), lub
nawet jeszcze wcześniej - w haplogrupie R1 (tamże?, Azja Zachodnia?), w której języki
satemowe hg R1a łączyły się, jako w jednym źródle, z kentumowymi językami hg R1b i
wszystkich jej podgrup (zob. niżej).
6.
Rozwój
europejskich Gałęzi Rodu
R1a. PraSŁOWIANIE
(Szczegółowo
o migracji i obecności R1a1a1 w rejonie Dunajca, Popradu i
Białej,
zob.)
 |
Ludność Europy (haplogrupy) na początku nowej ery
Haplogrupy ogólnie,
według
portalu
Eupedia.tree2.
Zob. rozmieszczenie R1a
na wschodzie, R1b na zachodzie,
I2a
na południu i wschodzie, I1, R1a, R1b na północy nordyckiej,
N na północy
rosyjsko-fińskiej i uralskiej,
E w
rejonie Grecji. |
Indoeuropejski ród R-M417 około 4.800
lat przed Chr. pojawił się w Europie środkowo-Wschodniej,
prawdopodobnie - według niektórych autorów - od Azji Mniejszej i
Bałkanów i na pewno jakoś uczestniczył
w
naddunajskich i środkowo-europejskich procesach rewolucji neolitycznej. W tym
czasie w skupionej populacji/rodu R1a1a musiał się dokonać proces
kształtowania się języka i etnosu, najpierw o wspólnym
charakterze indo-słowiańskim, a po rozdzieleniu się ludzi z
mutacjami Z93 i Z283 - oddzielonych etnosów prasłowiańskiego i
indo-aryjskiego.
Zob. umiejscowienie mutacji M417, A+, na
nici chromosomu Y: 8573735 kb (z FTDNA)
Z tego
też, zapewne naddunajskiego czy karpackioego terenu dokonały się
po około 3500 lat
przed Chr. migracje w różne rejony Europy, na zachód, północ i wschód. Czas i kierunek
wędrówek jest dziś rozpoznawany przez warianty haplotypów w STR w haplogrupie R1a1a1, zakodowane w Y-DNA dzisiejszych mieszkańców Europy
lub w znalezionych kopalnych szczątkach praprzodków, choć rzadko (bowiem
niektóre nowsze plemiona Prasłowian paliły swoich zmarłych; zresztą nie tylko
Prasłowianie). Etapy i kierunki migracji pozwalają też rozpoznać kolejne
mutacje SNP, aktualnie wykrywane w Europie i Azji, zwłaszcza
przez FTDNA.
Wynikiem
tych migracji był udział R1a1a1 w powstaniu rozmaitych kultur archeologicznych
nad Dunajem, zapewne np. ceramiki wstęgowej, oraz na północ i wschód od Karpat,
np. kręgu ceramiki sznurowej z kulturami grobów jednostkowych i
amfor kulistych.
http://en.wikipedia.org/wiki/Globular_Amphora_culture
Potem nad Dniestrem, Bohem i Dnieprem
dochodzi być może do symbiozy z
ostatnią fazą kultury Cucuteni-Trypolie
u wschodnich Karpat, od rzek Prut i Seret do Dniepru, a w
strefie północnej - z kulturą Bałtów - fatianowską.
Genealodzy genetyczni ustalili, że przodek
dzisiejszych wschodnich Słowian z hg R-M417 żył na ziemiach Ukrainy i Rosji
od około
2800-2600 lat przed Chr,
Następuje teraz rozwój kultury Prasłowian dalej w kierunku
wschodnim, ku Wołdze i Uralowi z jakimś udziałem w rozwoju kurhanowego
kręgu kultury jamowej (którą A. Klosov przypisuje populacji
R1b1a2). Na stepach pontyjsko-kaspijskich praindoeuropejskim kulturom przypisuje
się też udomowienie
konia, stosowanie powozu i upowszechnianie hodowli zwierząt
domowych (zob. Renfrew, Archeologia i język, PWN 2001, s. 249).
(Uwaga. Nie
znaleziono dotąd w rejonie Ukrainy i Rosji oczekiwanych przez
naukowców genetycznych śladów starszej ludności R1a1a, zwłaszcza
takich, których
by można datować na czasy ukraińskiego refugium podczas LGM,
którego domyślał się już
O.Semino i zesp. 2000,
tak jak je znaleziono na Bałkanach. Ukraina nie jest więc
pierwotną kolebką wszystkich Słowian. Zwłaszcza nie wolno ich
genezy datować tam na czasy niedługo przed V wiekiem po Chr., jak to głoszą
niektórzy archeolodzy polscy, tzw. skrajni allochtoniści ("dnieprowcy") np. K. Godłowski, a za nimi także niektórzy genetycy (zob.
Dyskusja n.5),
chyba wprowadzeni w błąd publikacjami znanego niemieckiego archeologa i rasisty Gustawa Kossiny (+1931). Około VI-VII w. po Chr., a więc
już w czasach historycznych, ówcześni Słowianie z Ukrainy swoją reemigracją
tylko może nieco zasilili osadnictwo południowych Słowian na Bałkanach, przerzedzone
klęskami w wojnach z cesarstwem wschodnim oraz na terenie Polski i krajów sąsiednich).
Dziś na Ukrainie, Białorusi i w Rosji żyje około 50% ludności
naszych haplogrup (w
niektórych starych miastach do 70%). Genealogia Y-DNA, prowadzona przez A.
Klosowa i współpracowników, zidentyfikowała nawet starożytnych założycieli dziewięciu
plemion ruskich z czasu około 2000 lat p.n.e i późniejszego, nazwanych imiennie dopiero we wczesnośredniowiecznych zabytkach
pisanych, m.in. bliscy nam Wołynianie, Bużanie i Biali Chorwaci
(kilka kopalnych szczątków ludzi owego osadnictwa hg R1a1a zidentyfikowano nawet w
Krasnojarsku w południowej Syberii).
Nieco
starsze niż na Ukrainie i
Rosji, bo z około 2800 lat p.n.e., korzenie plemion R1a1a1 znajdują się na
terenie Niemiec, a więc ród ów jest tam daleko
wcześniejszy niż geneza Germanów,
zwłaszcza tam, gdzie dziś nadal żyje skondensowane osadnictwo także słowiańskiej hg
R1a1a1g/g2 Serbołużyczan (choć zwykło się je
datować dopiero na wiek VII po Chr.!). Potwierdza
to np. niedawne, dokonane w 2005 r., słynne odkrycie i genetyczne
przebadanie cmentarzyska kilkunastu zmarłych,
a wśród nich trzech męskich osób z hg R1a1a w Eulau, 40 km za Lipskiem nad rzeką Soławą
(niem. Saale) w dorzeczy Łaby w Saksonii. Owe groby ofiar jakiejś
starożytnej przemocy pochodzą z około 2600 lat p.n.e i wyróżniają się świadectwami bogatej kultury życia rodzinnego i małżeństwa egzogamicznego.
Plemię to prezentowało kulturowe środowisko tzw. ceramiki sznurowej, kojarzonej
powszechnie
z Indoeuropejczykami. Archeologiczny haplotyp
mężczyzny z Eulau genetycy kojarzą obecnie ze staroeuropejską
lub
skandynawską gałęzią, wydzieloną wcześniej z ogólnego pnia.
Godni uwagi
są Nadłabscy Serbowie. jest możliwe, że w jakimś sensie oni, zanotowani przez tzw.
geografa bawarskiego jako Zeriuani (Serbianie?), byli tymi, "z
których wszystkie plemiona słowiańskie powstały i ród swój - jak
zapewniają - wywodzą". Wśród ludności niemieckiej do dziś około 10% wykazuje haplogrupę R1a1a1,
których wspólny przodek żył tam już może około 2700 lat p.n.e. Wyizolowani w
niemieckiej społeczności Serbołużyczanie słyną obecnie z tego,
ze jest u nich największe w Europie zagęszczenie hg. R1a1a1
(63%). Ale chyba niesłusznie, gdyż jeszcze większe zgęszczenie zostało
zauważone na Polskim Spiszu (73%,
zob.).
Z rodem R1a1a1 można ośmielić się kojarzyć powstanie i rozwój
środkowoeuropejskiej kultury amfor kulistych (http://en.wikipedia.org/wiki/Globular_Amphora_culture),
istniejącej w czasie 3400-2800 przed Chr. z centrum na terenie
Polski, która około 2900 lat przed Chr. Przeobraziła się ona w
rozległą kulturę ceramiki sznurowej, w tym kulturę grobów
jednostkowych i toporów gładzonych (http://en.wikipedia.org/wiki/Corded_Ware_culture). Jej południowy
region rozciągał się od Małopolski przez Śląsk i Czechy po Solawę (dopływ Łaby), z Serbołużyckim Eulau. Północny region tej kultury
przylegał do Bałtyku i rozciągał się od Kujaw i dolnej
Wisły, przez Wielkopolskę, rejon dolnej Odry i Meklemburgię po
Dolną Łabę (z tym horyzontem można kojarzyć późniejsze R1a1a w
jaskini Lichtensteinhohle z ok. 1000 przed Chr.), zob.
Dyskusja, n.16. Kultury
amfor kulistych i ceramiki sznurowej powszechnie uważane są za
pierwsze indoeuropejskie kultury w Europie. Także słynna kultura łużycka może być
w znacznym stopniu przypisana prasłowiańskiemu rodowi R1a1a1.
Populacje haplogrupy R1a1a1 i jej
podgrupy żyją dziś
w Polsce w największym w całej Europie jednorodnym zagęszczeniu, bo około 57%
ludności Polski.
|
Eupedia.tabl |
Na tablicy rozmieszczenie haplogrup w
dzisiejszych krajach europejskich; zob. R1a1a1 w czwartej
kolumnie.
Liczby oznaczają procenty owej haplogrupy w
poszczególnych krajach. R1b - to zachodnioeuropejska.
Zobacz skład etniczny (ojcowski, plemienny) ludności
dzisiejszej Polski (Poland) i jej sąsiadów |
Podobnie jak
w Polsce,
R1a1a1 (bez uwzględnienia podziału na jej podgrupy) pojawia się w Czechach (dziś 30%),
w Słowacji (40%),
na Węgrzech
(32%; poprzednia błędna liczba: 50%), w
Austrii - dziś ok. 25% i w Anglii (5% mieszkańców),
Irlandii (dziś m.in. klany Donaldów
i Douglasów), Szkocji;
Norwegii (do 30%
mieszkańców części zachodniej). Stąd właśnie osadnictwo R1a1a1 emigrowało
na Wyspy Brytyjskie i do
Islandii (dziś tam 23% ludności).
Nadto na Litwie,
Łotwie i Estonii dziś odpowiednio 38,
40 i 32%.
W sumie, na mapie geno-geografii
Europy największe zagęszczenie słowiańskiej oraz "byłej słowiańskiej" hg R1a1a1 ciągnie się szerokim pasem
od Bałtyku po Morze Czarne (co jest wodą na młyn tym, którym marzy się Polska "od morza
do morza"!).
|
W roku 2011, dzięki
badaniom Projektu 1000-Genomów, zidentyfikowano wiele nowych
mutacji SNP, określanych jako grupa Z albo "zetki".
Jeszcze nie
wszystkie z nich dostępne są w powszechnym testowaniu.
Najważniejsza z tych mutacji to
wspomniana wyżej Z93, cechującą
głównie azjatycką część rodu R1a1a1-M417.
Druga mutacja to
Z283; jej powstanie można datować
na około 3500 przed Chr. W tym rodzie zaistniały trzy ważne
mutacje:
Z280, cechująca głownie wielką
grupę, określaną jako Bałto-Slowianie. Jej synowską,
północno-wschodnią mutacja jest Z92.
M458
i L260 - cechujące ród
środkowoeuropejski i ród zachodniosłowiański (zwany tu polskim).
Trzecia z tych mutacji to
Z284, od której pochodzą
skandynawskie (i wyspiarskie) populacje rodu R1a.
Odkrywanie kolejnych mutacji i
testowanie jest w toku.
|
Procentowe rozmieszczenie czterech podstawowych mutacji (rodów) haplogrupy R1a w Europie
- według diagramu Igora Rożanskiego (stan
badań 2013 r.) (źródło:
http://www.rodstvo.ru/forum/index.php?showtopic=1312&st=740)
Dzieje wczesnych badań genealogii
genomicznej w hg. R1a
Problem pochodzenia Haplogrupy R1a, a zwłaszcza wielkiej
podgrupy R1a1a1-M417, jest jednak nadal dyskutowany. Według
niedawnej pracy zespołu hinduskich badaczy (np. ostatnio S.Sharma i
zesp., styczeń 2009) uważana do niedawna za ojcowską dla R1a1a haplogrupa R1a1-SRY1532.2
(faktycznie jednak R1a-M420) miałaby
się wyłonić w północnych Indiach na terenie Kaszmiru; albo
w środkowych Indiach (w plemieniu Saharia) około 18.400 lat temu (licząc 25 lat na
pokolenie). Tam wśród plemion Kashmiri Pandits zidentyfikowano 9
osób (4% spośród badanych) z tą haplogrupą, a w plemieniu Madhya Pradesh
Saharia 13 osób (23% badanych). Według tych badań, na terenie Kaszmiru lub Indii miałaby powstać także następna mutacja,
M17 i M198, dająca początek haplogrupie R1a1a.
Miałaby ona
zarazem dać - według tychże autorów -
początek hinduskim Indoeuropejczykom, głównej populacji Indii. Od roku 2010 wiadomo, że
mutacja R1a1a1-M417 jest najmłodszą
wspólną dla Europy i Indii. Obecnie do indoeuropejskiej populacji
R1a1a w Indiach należy do 17%
hinduskiego społeczeństwa (ponad 100 milionów ludzi). I co najciekawsze, ta głównie populacji
utworzyła najwyższą kastę hinduską, kapłańskich braminów albo
przynajmniej nią liczebnie zawładnęła . Przeciętnie blisko połowa, a w głównej ich
grupie - nawet 72% kapłanów-braminów wyróżnia się hg R1a1a, podkreślają badacze
hinduscy. Daty jakoby sugerują, że z hinduskiej populacji R1a1a
miałyby wywodzić się późniejsze populacje Słowian i innych
pokrewnych ludów. Autorzy zamieszczają daty tej haplogrupy na
Bliskim Wschodzie i w Europie - 11.200 lat temu, a w Azji Centralnej -
ok. 8600 lat temu.
Jednak
A. KLOSOW, częściowo
wspierany badaniami innych genetyków, wskazał "kolebkę" R1a1a
ogólnie w południowej Syberii, w pobliżu Chin. Według Klosowa powstanie mutacji
M420, a zatem i haplogrupy R1a, dokonało się gdzieś w okolicach gór Ałtaju, gdzie wśród niektórych
plemion, dziś językowo ałtajskich, tureckich i rosyjskich, występuje ojcowska haplogrupa
R1-M173 do 20%, czyli w stosunkowo najwyższym zagęszczeniu;
choć występuje ona także w Azji Środkowej i na Bliskim Wschodzie
- w mniejszym procencie).
Oto przytaczane przez niego
fakty:
Anatole Klyosov 2009
w piśmie Вестник Российской Академии
ДНК-генеалогии
(Wiadomości Rosyjskiej Akademii genealogii Y-DNA) Том 2, № 5 2009 август,
w artykule
Древнейшие
восточно-азиатские ветви гаплогруппы R1a,
(Stare wschodnioazjatyckie gałęzi haplogrupy R1a) dokonał obliczeń i ustalił, że
R1a1a wywodzi się z południowej Syberii; mutacja M17
(i M198) powstała
tam 20.000 lat temu
bowiem na taki czas wskazują jej haplotypy STR,
które Klosov w swojej pracy opublikuje.
Doszedł do wniosku, że jedna część tej
populacji z południowej Syberii wyemigrowała przez
północnowschodnie i środkowe Chiny do południowych Indii
i Pakistanu; w Indiach stanowi starą populację wśród Drawidów, ze wspólnymi przodkami z czasu
7000-12000 lat temu.
Druga część, która pozostała "na miejscu" w Syberii, dała
później początek być może niektórym starym
tamtejszym kulturom archeologicznym, jak np. taszbułatowskiej (8000-7000 lat) i
Afanasjewo (rejon Ałtaju 4500-4000 lat temu).
Trzecia zaś część wyemigrowała na zachód, docierając
około 12.000 lat temu na Bałkany, by po kilku
tysiącach lat około 5000-4000 rozejść się po Europie, w
tym także do pontyjsko-kaspijskich stepów, oraz dalej
na wschód, za Ural, tworząc kulturę Sintaszta i Andronowo,
gdzie uformowała sie populacja Ariów, która w północnych
Indiach dała początek młodszej, indoeuropejskiej części
R1a1a (o czym niżej).
Datowanie haplogrupy R1a1a na czas około 20.000 lat
pociągnie za sobą wcześniejsze datowanie powstania
także mutacji M173, czyli haplogrupy R1, datowanej przez
T. Karafet et al. 2008 na czas 18.500lat temu.
Klosow jednak dodaje, że konieczne są
dalsze ustalenia w temacie datowania i migracji hg. R1 i R1a1a oraz
jej rozprzestrzeniania się.
Najnowsza publikacja o
starych azjatyckich haplogrupach, w tym R1a1a, zob.
Shou et al. 2010 Ocena artykułu w
Rodstwo.ru
Klosow,
przeprowadzając rewizję dotychczasowego datowania haplogrup europejskich i
azjatyckich, w tym także R1a1a (R1a1a1), zauważył, że dokonane przez historyków,
językoznawców i archeologów datowanie inwazji półwyspu Indyjskiego przez Ariów z
północy i zaistnienie tam indoeuropejskiego języka dopiero na połowę drugiego tysiąclecia p.n.e. jest zgodne
tylko z poprawnym datowaniem
i genealogią hinduskiego haplotypu R1a1a, wyraźnie identycznego z haplotypem
wschodniosłowiańskim, lecz nieco młodszego.
Bowiem spośród
Prasłowian wschodnich, których wspólny przodek wywędrował około 2750 lat p.n.e na ukraińsko-rosyjskie stepy, część populacji migrowała dalej na wschód, na teren
dzisiejszego północnego Kazachstanu i południowego Uralu, tworząc tam rozległą kulturę andronowską, z centrum nad Jeziorem Aralskim.
Prawdopodobnie z tej populacji niewielka społeczność zbudowała
sobie około
1800 r. p.n.e m.in. gród/zamek o nazwie Arkaim na południowym krańcu Uralu.
Został on jednak porzucony po około 200 latach. Bowiem po
jakiejś katastrofie, którą można kojarzyć z wybuchem
około 1628 lat p.n.e wulkanu na
wyspie Thera (dzisiejszy jej fragment to Santorini) na Morzu Egejskim
i
który pochłonął cywilizacje minojską, a na szerokiej przestrzeni północnej
półkuli na długi czas zakłócił pogodę (znaczne ciemności i obniżenie
temperatury) ludność Arkaim z R1a1a1 porzuciła gród i udała sie na południe,
zaludniając Kirgizję i Tadżykistan (tam dziś
wśród ludności - do kilkadziesiąt procent genetycznych ex-Słowian R1a1a1!) oraz
północne Indie, a chyba osobno - północny Iran (badania irańskie są dotąd
niewystarczające).
Co do hinduskiej populacji R1a1a1 trzeba wiedzieć i brać pod
uwagę, że na podstawie tekstów hinduskiej
świętej księgi Rygwedy i innych badań, zwłaszcza archeologii i językoznawstwa,
światowa nauka dotąd uważa indoeuropejski lud Ariów, który dał początek kastom
braminów hinduskich i hinduskim Indoeuropejczykom, raczej za najeźdźców z
północy w połowie II
tysiąclecia p.n.e., co potwierdza genealogia genetyczna. Dlatego Genetyka na
portalu Eupedia podkreśla: "Kapłańska kasta
braminów składa się niemal wyłącznie z haplogrup R1a1, R2
i J2a (chociaż R1a1a stanowi dwie trzecie z linii), przybyłych w
ramach indoaryjskiej migracji z zewnątrz w epoce brązu 3500 lat
temu". Genealogia Y-DNA w badaniach A.K.
zdaje się potwierdzać pochodzenie językowo indoeuropejskich Ariów od
Prasłowian wschodnich, o czym niżej, w rozdziale o ekspansji
Prasłowian.
Obecnie
[2012 rok] już wiadomo, że gdzieś w rejonie
południowego Uralu, Morza Kaspijskiego lub okolic Jeziora Aralskiego doszło do
powstania prężnej populacji z mutacją
Z93 i jej synowską L342.2, wykrytą
w lipcu 2011 r. Grupa ta żyła zapewne na uboczu od reszty rodu
R1a1a1, gdyż potem, pod nazwą
Ariowie, bez przemieszania się z bratnimi
Prasłowiańskimi mutacjami spod haplogrupy Z283, migrowała na południe, w
kierunku Indii i Iranu, oraz na Wschód, wraz ze swoimi językami z grupy
indoeuropejskiej. Mutacje Z93 i L342.2 odróżniają populacje
azjatyckie od Europejskich, uważanych do niedawna przez
rosyjskich genealogów za nieodróżnialne.
1. I. Rożanskij i A. Klosow, listopad 2009
- kompendium aktualnej
wiedzy o europejskiej części populacji R1a,
(gałęzie przed
rozpoznaniem mutacji M458 i haplogrupy R1a1a7)
"Haplogrupa R1a: haplotypy, linie genealogiczne, historia,
geografia"
И. Рожанский и А. Клёсов, Гаплогруппа R1a: гаплотипы, генеалогические линии, история,
география,
w:
Вестник Российской Академии ДНК-генеалогии
t.2, nr.6 (listopad), 2009 r., s. 974-1099.
2. A. Klosow i I. Rożanskij, grudzień 2009
o haplogrupie R1a1a1g, czyli gałęzi
środkowoeuropejskiej i zachodniosłowiańskiej
"Podgrupa R1a1a1g - M458 - populacje, geografia, historia"
w:
Вестник Российской Академии ДНК-генеалогии
t.2, nr 7 (grudzień), 2009 r., s.1200-1216.
w języku rosyjskim, gruntowny komentarz do pracy Underhilla et
al. 2009 (zob. niżej) w temacie haplogrupy R1a1a1g-M458,
uzupełnienie pracy Rożanskiego-Klosowa
"Haplogrupa R1a: haplotypy, linie genealogiczne, historia,
geografia"
Вестник
t.2, nr 6, 2009 r. (zob. wyżej).
3. I. Rożanskij i A. Klosow, styczeń 2010
o bliższym genealogicznym związku gałęzi skandynawskich z
kirgiską.
"Migracje z południowej
Syberii i Azji Środkowej z punktu widzenia genealogii Y-DNA"
Вестник Российской Академии ДНК-генеалогии,
tom
3, nr 1, 2010 r.
4. I. Rożanskij - 2009-2010
kilkanaście
map rozmieszczenia gałęzi haplotypów R1a1a1
Google-mapy;
5. P. Szwarew, Forum Rodstvo.ru,
omówienie map i drzew genealogicznych
Гаплогруппа R1a: гаплотипы, генеалогические линии,
история, география, И. Рожанский и А. Клёсов
6. I Rożanskij, A. Klosow,
styczeń 2010
"Migracje z południowej Syberii i Azji Środkowej do
północnej Europy z punktu widzenia genealogii Y-DNA"
Миграции из южной Сибири и Средней Азии в северную
Европу с точки зрения ДНК-генеалогии
w: Вестник Российской Академии ДНК-генеалогии Том 3, № 1
2010 январь
http://www.lulu.com/items/volume_67/8049000/8049755/2/print/8049755.pdf
Prace rosyjskich genealogów Y-DNA
Analizą
mutacji SNP i STR, czyli haplogrup i haplotypów, oraz
budową drzewa filogenetycznego i opisywaniem jego gałęzi zajmują
się głównie rosyjsko-amerykańscy
naukowcy.
I. ROŻANSKI i A. KLOSOW w swoich
obszernych publikacjach dokonują uporządkowania i
genealogicznego opracowania obecnej wiedzy o populacjach haplogrupy
R1a i jej podgrup oraz ich historii w Europie
i Eurazji. Autorzy mają aktualnie do dyspozycji ponad 2000 haplotypów z bazy Ysearch
i geograficznych projektów FTDNA oraz z laboratoriów rosyjskich
i prywatnych; najwięcej 67-markerowych.
Praca P. Underhilla (Uniwesytet Stanford)
Publikacja
Underhill et al. (publikacja
4.11.2009), znacznie zmieniła dane wcześniejszych autorów,
wskazały na okolice Polski jako na praojczyznę Słowian i
miejsce powstanie polskiej mutacji R-M458 dla
słowiańskiej populacji Europy. Oto niektóre
dane polskie i wybranych krajów.
Czas wspólnego
przodka podano w tysiącach lat temu, obliczony jest według
stawek Zhivotovsky'ego, a więc chyba niewłaściwie obliczonych i dwu-trzykrotnie zawyżonych.
 |
Na lewo: mapa
zagęszczenia, czasu haplogrupy R1a1a (M17) i drzewo
genealogiczne grupy i podgrup.
Na prawo: mapa zagęszczenia, datowania i związku
z kulturami archeologicznymi populacji R1a1a1g (M458) -
według: Underhill et al. 2009. |
 |
http://www.scribd.com/doc/23322591/Underhill-Et-Al-2009-
Separating-the-Post-Glacial-Coancestry-of-European-and-Asian-Y-Chromosomes-Within-Hap-Lo-Group-R1a
Oto kilka danych Underhilla odnośnie
(niestety zawyżonego) datowania populacji R1a1a-M17 i R1a1a1g -M458 w kilku
krajach: Polska
- lat 11300 i 10700; Słowacja 11200 i 8300;
Czechy - 5700;
Niemcy 9900 i 7500.
Rosja europejska 8700 i 8000;
Ukraina 7400 i 4700;
Indie 14000;
Pakistan 15000.
Późniejsze wykrycie w tej
słowiańskiej podgrupie kolejnej mutacji, oznaczonej jako
L260 (R1a1a1g2) i nazwanej P ("polska") jeszcze bardziej
związało początki Słowian z tym rejonem Europy.
Prace polsko-amerykańskich genealogów P. Gwozdzia i
L. Mayki
W tych pracach Rosjanom dotrzymują
kroku, a niekiedy przewodzą polsko-amerykańscy genealodzy -
współpracownicy FTDNA: Peter
GWOZDZ (USA) i Lawrence
MAYKA (USA), którzy zajmują się genealogią
wszystkich haplogrup (a nie tylko R1a), ale jakoś
związanych z Polską
* * * *
Najstarsze haplotypy z haplogrupy R1a w Europie zostały w 2008
r. rozpoznane przez A. Klosowa wśród ludności
niektórych krajów bałkańskich: Serbia, Bośnia, Kosowo i
Macedonia, w materiale, który zebrali i opublikowali M.
Perićić i L.Barać. A. Klosow datuje je na około
dziesiąte tysiąclecie przed Chr. Niesprzyjające
warunki bytowe (tzw. szyjka butelki) lub dryf populacji
zdziesiątkowały ich liczebność i nie pozwoliły im na
pełny rozwój demograficzny. Zanim jednak do tego kryzysu
doszło, około polowy trzeciego
tysiąclecia przed Chr., owe populacje haplogrup R1a1a miały zapewne wydatny wpływ na
rozwój archeologicznych kultur naddunajskich,
środkowoeuropejskich i dalszych. Archeogenetyka zapewne
będzie miała szansę wypowiedzieć się na ten temat, jak
to się stało już odnośnie szkieletów z Eulau,
Lichtenstein-Hohle, Krasnojarsk itd.
Zdaniem jednak wielu genealogów genetycznych ów materiał
"bałkański", choć zebrany przez solidne ośrodki naukowe,
nie jest w pełni wiarygodny.
Zespół genetyków
P. Underhill et al. (listopad 2009, zob. niżej) wykrył zaś ważną
europejską mutację R-M458, tworzącą nową haplogrupę
R1a1a1g, datując ją, z grubą przesadą, na dziewiąte
tysiąclecie przed Chr. i (dość trafnie) wskazując na
rejon Polski jakby na
teren jej powstania i jakby na praojczyznę
znacznej populacji Słowian środkowoeuropejskich,
zwłaszcza polskich. Kolejno, po wykryciu w haplogrupie R1a1a1g
mutacji L260 ustalono haplogrupę R1a1a1g2, która
definiuje wśród Słowian gałąź "polską", a
raczej zachodniosłowiańską.
Zaś szczególną zasługą
genealoga genetycznego L. Mayki (USA) jest
zidentyfikowanie i zorganizowanie testowania ważnej
mutacji "azjatyckiej", czyli L342.2, która w olbrzymim rodzie
R1a1a1(M417) wydzieliła populacje środkowo i
południowoazjatyckie oraz żydów-lewitów aszkenazyjskich.
AKTUALNE PRACE
GENEALOGII GENETYCZNEJ
L. Mayka prowadzi program
FTDNA Polish Project . Rejestruje tam się wyniki
analiz przeprowadzonych w pracowniach FamilyTreeDNA
dotyczących materiałów z DNA, dostarczonych przez
Polaków;
internetowa strona przedstawia rezultaty badań i identyfikacje haplogrup
Polaków, a w haplogrupie R1a1a1g - podział na klastry, w zależności od haplotypów STR.
Dokładnym rozpracowaniem
polskich haplotypów z haplogrup R1a1a i R1a1a1g zajmuje
się P. Gwozdz. Na swojej internetowej stronie
Polish clades prowadzi analizę haplotypów, grupuje, oblicza
czas populacji i ich migracji. W opublikowanej w JOGG
Journal of Gnetic Genealog 5/2 2009 jego pracy i pojawia się więcej informacji
na temat metody pracy i jej rezultatów (część II).
Czas wspólnego przodka haplotypu P .
Prace zespołu Łukasza Łapińskiego.
Całością haplogrupy R1a jej
mutacji i
podgrup w świecie oraz podziałem na genetyczne gałęzie
zajmuje się od czerwca 2011 r. genealog Łukasz Łapiński
na stronie
R1a and Subclades Y-DNA Project.
(Natomiast podziałem tego rodu na
poszczególne kraje lub większe grupy narodowościowe i
regionalne zajmuje się strona Cyndi (Kanada):
R1a Y-chromosome Haplogroup Project.)
|
GENEZA JĘZYKÓW SATEMOWYCH (indo-bałto-słowiańskich)
Na ojczystym terenie, zapewne gdzieś w rejonie Dunaju
lub Karpat, w populacji R1a1a1 dokonała
się satemizacja języka (dotąd zapewne kentumowego
jak nadal w haplogrupie R1b), równoznaczna z
formowaniem się języka praindo-prabałto-prasłowiańskiego.
Satemizacja polegała głównie na innowacji, że pierwotnie miękkie
k' u satemowców przeszło w s (u kentumowców
tylko utraciło
zmiękczenie), stając sie zwykłym k lub c. Wnet potem,
od około III tysiąclecia p.n.e. (ale jeszcze przed
terytorialną ekspansją Prasłowian),
nastąpiło oddzielenie
się języka
indo-irańskich Ariów (głównie z mutacją Z93), a
później oddzielenie się
języka bałtyjskiego.
Ostatecznie rodowi z mutacją
Z283, żyjącemu na na niewielkim terenie Europy,
zapewne
w Kotlinie Dunajskiej lub w
rejonie Karpat, należy
przypisać dokonanie podstawowych prasłowiańskich
innowacji językowych, których zaistnienie dopiero po
rozprzestrzenieniu się na wschód i rozproszeniu ludności
byłoby już niemożliwe (por.
I. Rożanskij i A. Klosow,
Wiestnik, t.2, n.6) |
UWAGA.
W rejonie Europy Środkowej
nastąpiło spotkanie ludności hg. R1a1a z ludnością
kultury pucharów lejkowatych (zapewne hg. I1 i I2), oraz zainicjowanie i/lub upowszechnianie
kilku innych kultur, zwłaszcza kultury amfor kulistych i ceramiki sznurowej (Eulau)
z toporami gładzonymi i grobami jednostkowymi. Regres
demograficzny, trwający w drugim
tysiącleciu przed Chr. w rejonie Europy Środkowej, nie
był całkowity i nie oznaczał katastrofy
kulturowej, zwłaszcza że na sąsiednich ziemiach
dzisiejszej Ukrainy i Rosji Europejskiej tego regresu
nie było, co umożliwiło późniejsze wsparcie
demograficzne i kulturowe dla ziem rejonu Środkowej
Europy.
Postscriptum.
Powyższe badania zdają się nie potwierdzać
tezy niektórych archeologów o naddnieprzańskim pochodzeniu
Polaków i ich haplogrupy R1a, i to dopiero w V wieku po Chr. Co więcej,
wyraźnie zauważa się, że Polska leży jakby w centrum
starożytnego osadnictwa słowiańskiego haplogrupy R1a. W Rosji np. o rezultatach
pobierania próbek DNA wprost pisze się: Poskrob
Ruskiego - znajdziesz Polaka (
Поскреби русского - найдешь поляка,
Газета.ru
14.01.2008) - jak przedtem, podobno niesłusznie, tam się mawiało:
Поскребешь русского
- найдешь татарина - 'Podrapiesz Ruskiego, znajdziesz Tatara'.
* * * * *
Haplotypy JTJ.pdf
Archeologiczne (ancient)
Y-DNA w Europie.
Tabela i ilustracja zasiedlenia starożytnej
Europy według znalezisk archeologicznych Y-DNA;
w kulturach, czasie i procentach (stan badań
grudzień 2015)
(źródło: Eupedia, Tomenable/Davidski)
http://s14.postimg.org/mu6jakwv5/Chronological_YDNA.png
http://www.tropie.tarnow.opoka.org.pl/images/ancient.ydna.eur.png

(klik-powiększenie)
Adres:
ks. Stanisław Pietrzak,
Tropie 6,
33-316 Rożnów
E-mail:
pietrzakstan@poczta.onet.pl
Wszelkie uwagi dla mnie proszę
przekazać meilowemu gołąbkowi:

Dziękuję,
już dziś przekażę Adresatowi
|
DZIĘKUJĘ!
JESTEŚ MIŁYM |
 |
GOŚCIEM
NASZEJ WITRYNY!
|
Do strony Pietrzakowie. Genealogia.
Pietrzak
Do strony Kurczabowie. Genealogia.
Kurczab. Bołoz
Do strony Janikowie. Genealogia. Janik
Do strony Szlagowie. Genealogia.
Szlaga/Ślaga
Powrót do Strony
głównej witryny: Parafia TROPIE
http://ymap.ftdna.com/cgi-bin/gb2/gbrowse_details/hs_chrY?name=L74
Str.pryw.-genealog.
PIETRZAKOWIE.Pietrzak,
JANIKOWIE.Janik,
FIUTOWIE,
SZLAGOWIE Nadto:
Gierałt.GIERAŁTOWIE
*
|
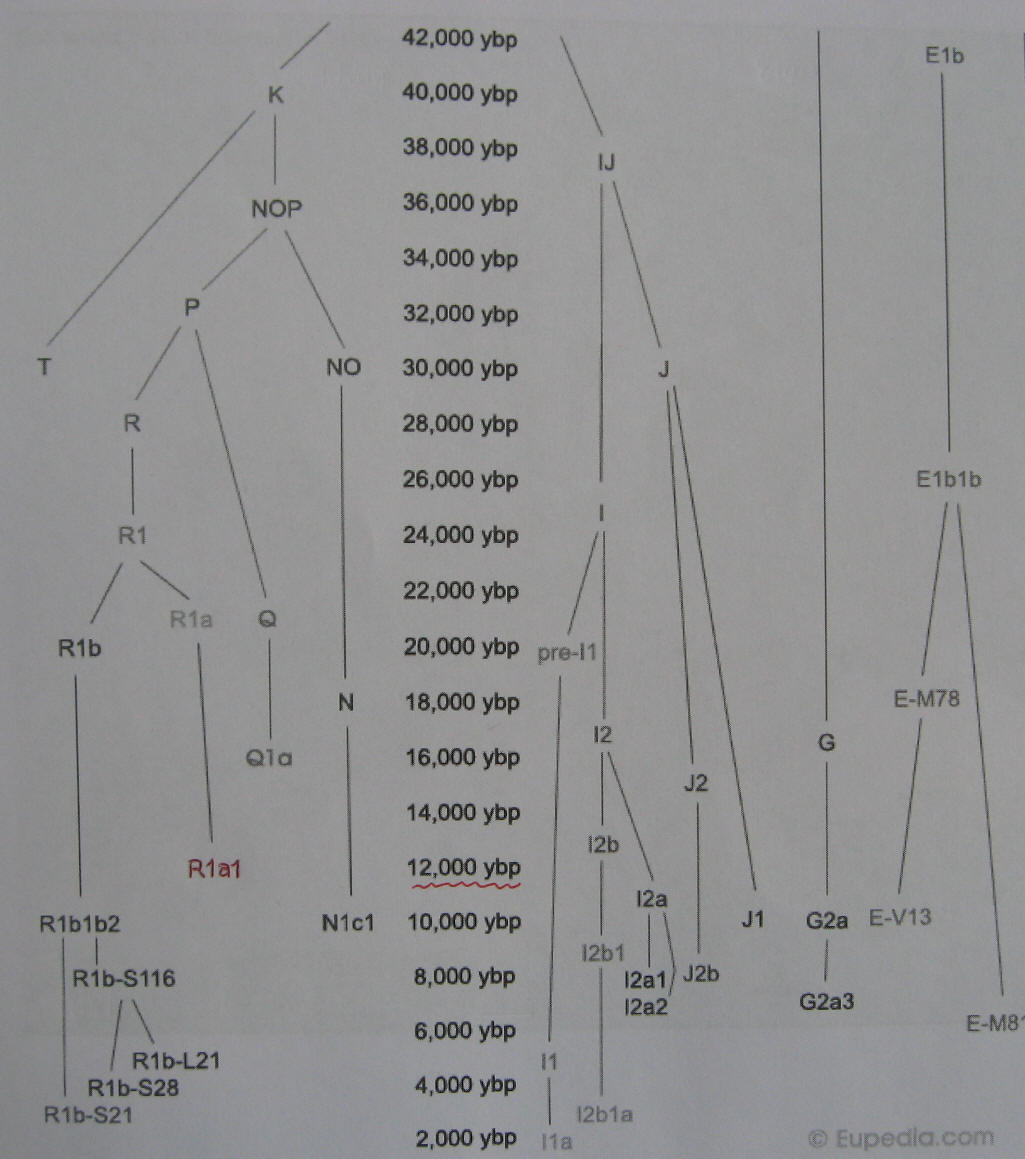


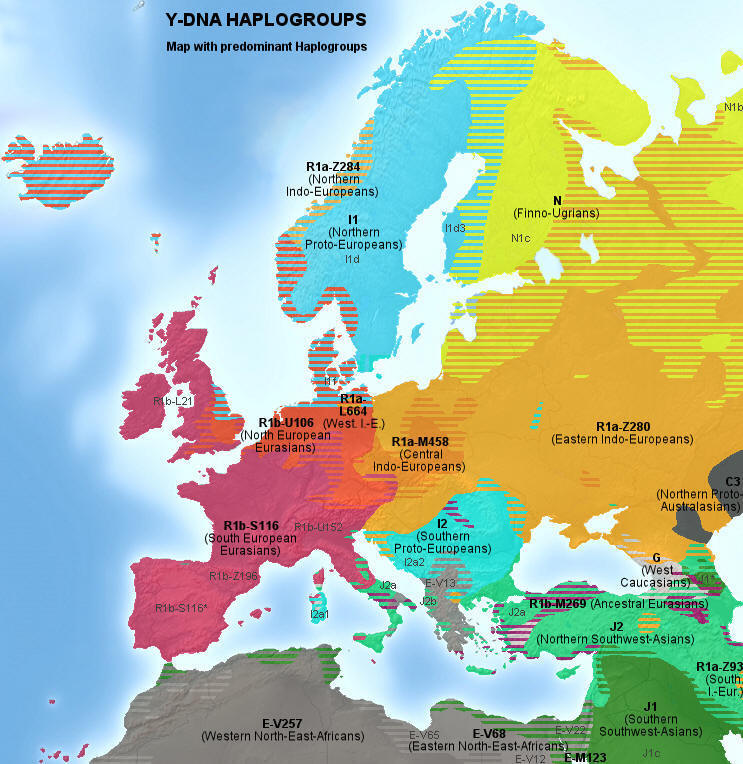
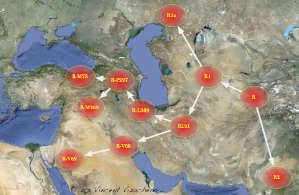
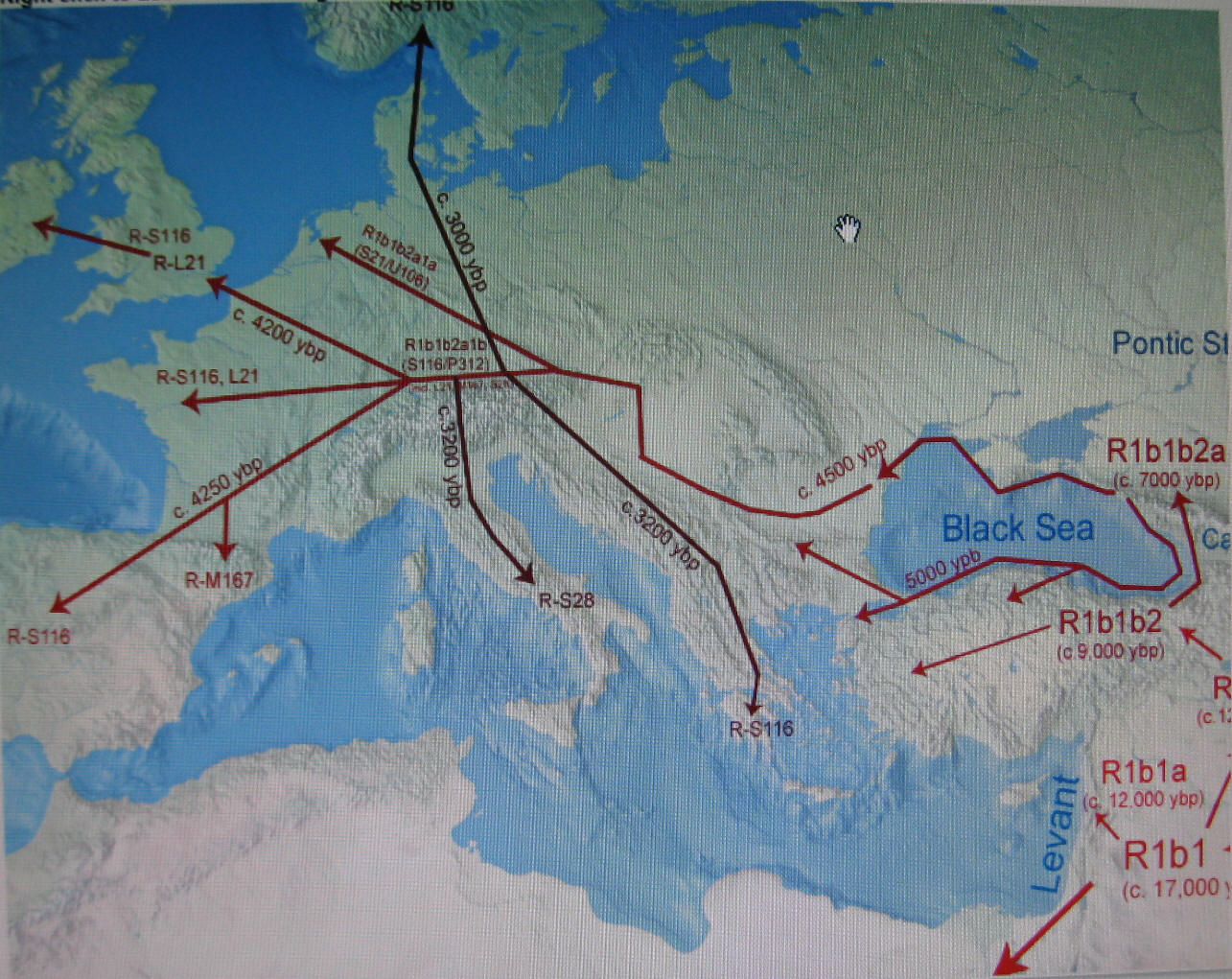
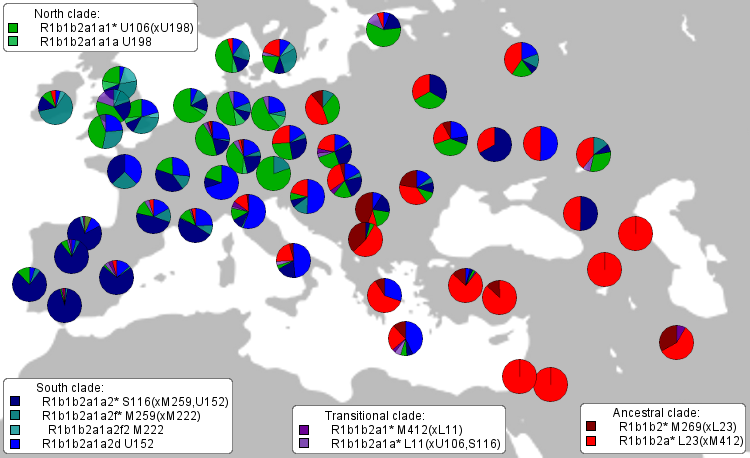
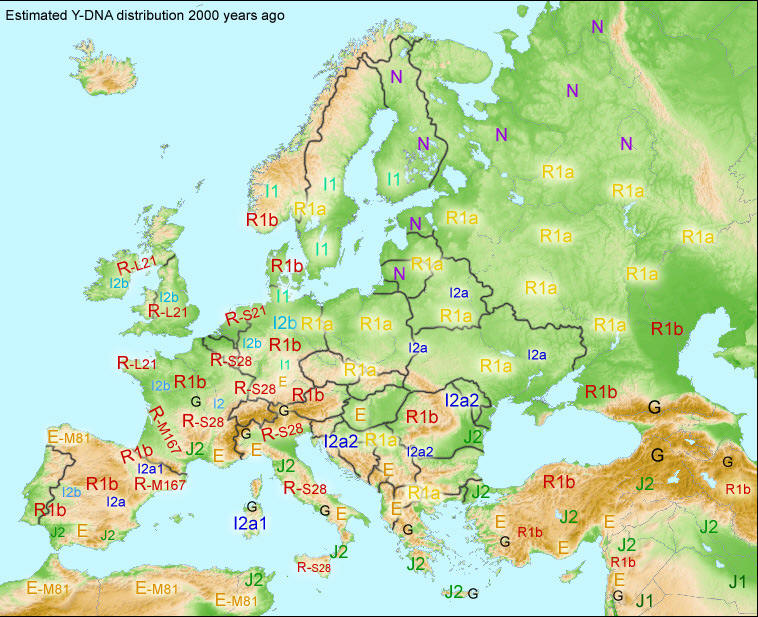
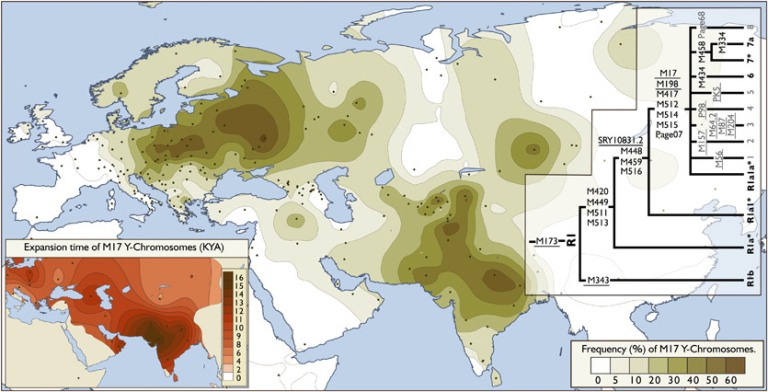

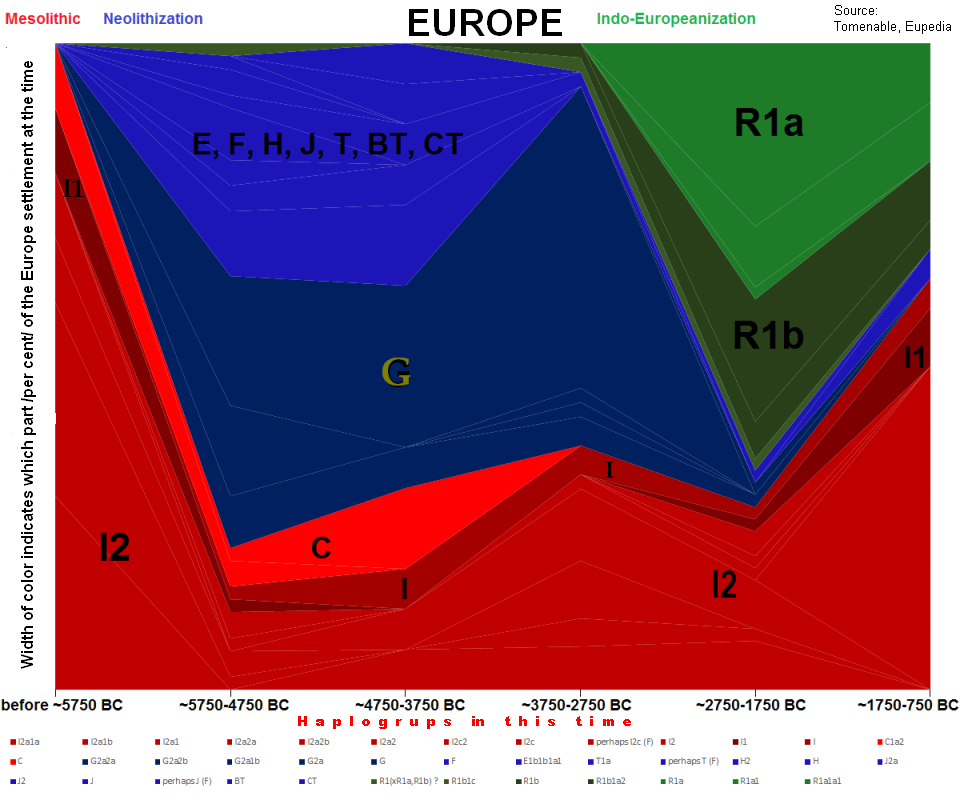
{kind=link}
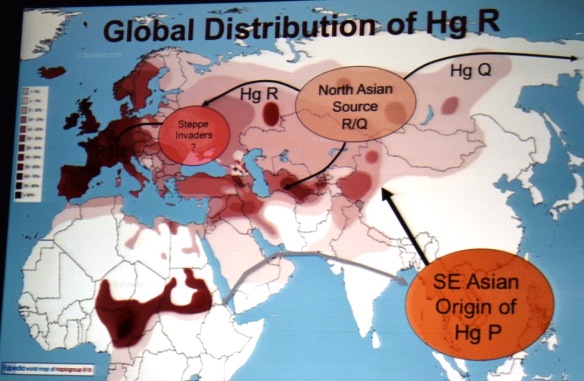{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}