|
Wiele tysięcy praojców L-74, R1a-YP380, mt-H14a |
Genetyka odsłania tajemnice:
Pochodzenie ludności
|
Ostatnia
aktualizacja |
|
†
.
Genetyka odsłania
tajemnice
pochodzenia
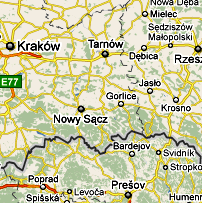
Lokalne, regionalne i ponadregionalne periodyki, poświęcone kulturze, historii i sprawom etnicznym, często starają się odpowiedzieć na frapujące pytanie o początek takiej, czy innej, omawianej rzeczywistości etnicznej. Odpowiedzi szukają zwykle w pamięci świadków, źródłach pisanych, pamiątkach rzeczowych lub nawet w archeologii. Ale te źródła w wielu sprawach okazują się ograniczone i niezdolne do udzielenia pełnej odpowiedzi na postawione pytanie. Do takich pytań należy i kwestia pochodzenia grup ludności w rejonie Dunajca, Popradu i Białej lub najbliższych terenów przyległych. Nauka przecież już nie pozwala nam odpowiadać: Jesteśmy tu “od zawsze”, bo przecież rodowym gniazdem człowieka współczesnego nie jest ani Polska, ani Europa, lecz środkowo-wschodnia Afryka. A jeżeli nie jesteśmy tu, w rejonie Dunajca, Popradu czy Białej “od zawsze”, to kiedy, skąd i jakimi szlakiem migracji przybyli tu nasi praojcowie? Czy przybyli z jednego, czy wielu regionów naszego kraju, Europy, Azji czy Afryki? Czy to prawda, że Słowianie pojawili się dopiero około V-VI wieku po Chrystusie i jakby z nikąd, co kiedyś starał się udowadaniać niemiecki archeolog-rasista Rudolf Kossina, a teraz także nasi skrajni allochtoniści?
I. Wyjaśnienie pojęć genealogii genetycznej,
Na postawione wyżej pytania stara się odpowiedzieć genetyka, a właściwie genealogia genetyczna zwana też genealogią Y-DNA, czyli płciowego chromosomu męskiego oraz genealogia mtDNA, czyli przekazywanego przez matki “energetycznego” genu w komórkach organizmu. Jest to nowa gałąź wiedzy, licząca zaledwie około dziesięciu lat. Źródłem dla tej nauki są dane z testów Y-DNA i mtDNA w laboratoriach, dotąd przeważnie amerykańskich albo zachodnioeuropejskich. Dla potrzeb genealogii analizuje się dwa rodzaje mutacji Y-DNA, które na podobieństwo archiwum historii przechowywane są w niekodującym (“śmieciowym”) regionie Y-DNA i przekazywane w liniach prostych tylko przez ojców i tylko swoim synom od pierwszego praojca, "chromosomalnego Adama” aż do dziś. Jedne mutacje, określane skrótem SNP (Single Nucleotide Polimorphism), zachodzą bardzo rzadko, zasadniczo raz w dziejach współczesnego człowieka i one wyznaczają niepowtarzalne haplogrupy SNP (od gr. haplos ‘pojedynczy, jedyny, niepowtarzalny’). Haplogrupy pozwalają odróżnić wielkie populacje ludzkie, liczące nawet dziesiątki lub setki milionów ludzi.
Inne mutacje, zachodzące w rejonach szybkozmiennych Y-DNA (np. co kilkadziesiąt lub kilka tysięcy lat), oznaczane skrótem STR (Short Tandem Repeats), wznaczają tzw. haplotypy STR indywidualnych ludzi. Haplotypy pomagają odróżnić ludzkie rody, rodziny lub niekiedy nawet poszczególne osoby w rodzinie. Porównanie kilku lub więcej haplotypów STR umożliwia zidentyfikowanie rodzaju i ilości mutacji oraz ustalenie wieku ich wspólnego przodka, określanego angielskim skrótem TMRCA (Time to Most Recent Common Ancestry). Analizuje się też geny mitochondrialne. Mitochondria znajduja się w plaźmie komórkowej i odpowiadają za jej energetykę (oddychanie i utlenianie w komorkach). Przekazywane są tylko w liniach prostych i tylko przez matki swoim dzieciom (córkom i synom). W mitochondriach też - jakby w bibliotece - “przechowuje się” seria mutacji w linii matczynej, począwszy od pierwszej matki, "mitochondrialnej Ewy”, aż do dziś. MtDNA testuje się w dwóch rejonach szybkozmiennych: HVR1 i HVR2, a w pełnym badaniu - także rejon pośredni między nimi. Dzięki tym mutacjom w Y-DNA i mtDNA możemy poznać swoją ojcowską czy matczyną genealogię w linii prostej od pierwszych prarodziców, szlaki ich migracji ku naszej małej ojczyźnie, a także rozpoznać stopnie pokrewieństwa z innymi populacjami ludzkimi (tzw. genetyczny dystans, oznaczany symbolem GD). W laboratoriach obecnie standardowo testuje się 67, a od kwietnia 2011 nawet 111 markerów, czyli ustalonych miejsc na “nici” Y-DNA. Takie analizy przeprowadza zwłaszcza amerykańska firma komercyjna Family Tree DNA (FTDNA) przy Uniwersytecie Arizona. W Rosji firma rosyjsko-czeska Gentis ogranicza się aktualnie do badania tylko 17 markerów (miejsc), co nie daje podstawy do dokładniejszych badań genealogii. Podobne badania, niekiedy bardzo szczegółowe, przeprowadzają też liczne zakłady medycyny sądowej, ale przeważnie według specjalnych wymagań sądownictwa i kryminalistyki. Wyniki testów swojego DNA klienci przekazują dobrowolnie do baz Y-DNA i mtDNA dla potrzeb innych badań naukowych, zwłaszcza genealogicznych. Największą bazę stanowi Ysearch, w której znajdują się wyniki testów już blisko setek tysięcy ludzi z całego świata; medycyna sądowa posiada odrębną, dla genealogów mniej przydatną bazę: Yhrd. Opracowania tych wyników publikowane są w międzynarodowych periodykach naukowych, dostępnych u nas w uprzywilejowanych bibliotekach uniwersyteckich lub także przez internet. Z punktu widzenia naszych polskich potrzeb najbardziej odpowiednie są dwa środowiska naukowe, opracowujące wyniki testów, jedno w USA, a drugie w Rosji. To środowisko w Stanach Zjednoczonych tworzą dwaj Amerykanie polskiego pochodzenia, genetycy Lawrence Mayka którego praojcowie pochodzą z Dębicy i Peter Gwozdz, którego przodkowie wywodzili się z okolicy Mielca. Pierwszy prowadzi tzw. FTDNA Polish Projekt, w którym zbiera i segreguje wyniki badań DNA ludzi, mających jakikolwiek rodowy związek z Polską w jej dawnych granicach. Ten drugi zajmuje się teoretycznymi badaniami grup i podgrup zebranego materiału polskiego, zwłaszcza jego datowaniem. Środowisko rosyjskich genetyków nie ogranicza się do danych z terenu Rosji, lecz analizuje cały zestaw danych, zwłaszcza z bazy Ysearch, ze szczególnym uwzględnieniem materiału z terytoriów słowiańskich. Jeden ośrodek tworzy laboratorium Gentis i jego strona internetowa MolGen. Drugi to ośrodek zwany Rosyjska Akademia Genealogii Y-DNA, którą kieruje amerykański uczony rosyjskiego pochodzenia, dr Anatol Klosow (Anatole A. Klyosov). Jego zainteresowania koncentrują się także na tworzeniu jakby światowej wspólnoty duchowej genetycznego rodu R1a1a1 (rodu głównie słowiańskiego). Od czerwca 2011 r. inicjatywę w opracowywaniu danych całej haplogrupy R1a w świecie (i tylko tej) przejęła międzynarodowa grupa wybitnych genealogów genetycznych, zorganizowana przez polskich badaczy pod kierownictwem dra Łukasza Łapińskiego. Na portalu R1a1a and subclades Y-DNA project grupa stymuluje rozwój testowania, analizuje haplotypy, śledzi nowe SNP (t.j. snipy), sporządza cenne mapy rozprzestrzenienia się poszczególnych rodów i ich podgrup. Grupa zaznacza też swoją obecność na forum dyskusyjnym A Genetic Genealogy Community dla przekazywania informacji i kontaków z klientami. Grupa oficjalnie używa języka angielskiego, a nieoficjalnie polskiego i nadto kilku innych. Światowe stowarzyszenie genetyków (YCC) zadecydowało oznaczać niektóre mutacje SNP w Y-DNA, wyróżniające przodków wielkich populacji ludzkich, kolejnymi dużymi literami alfabetu od A do R lub dalej. Większość z nich należy do pnia drzewa genealogicznego ludzkości. Po zmodyfikowaniu pierwotnych oznaczeń pień drzewa tworzą haplogrupy: A, A1 BT, CF, F, IJK, K', K'', (NO), P i R. Natomiast inne duże i małe litery oraz cyfry oznaczają dalsze podporządkowane mutacje SNP. W ten sposób np. w Polsce i sąsiednich krajach słowiańskich najczęściej występuje przynależność do haplogrupy R1a, a zwłaszcza wielkiej podgrupy R1a1a1, którą wyznaczyła mutacja M417 (litery M, L, P, CTS itd są oznaczeniami laboratorium, które wykryło daną mutację, czyli SNP)).
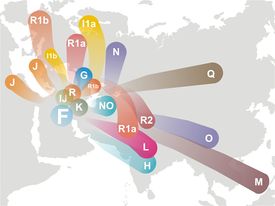
Zobacz drzewo genealogiczne
haplogrup/rodów Y-DNA, według Eupedii:
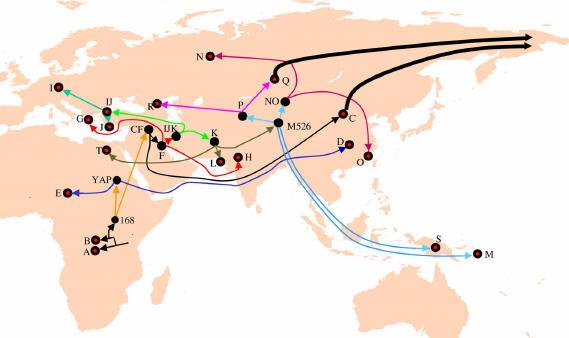
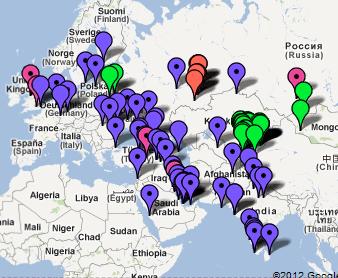
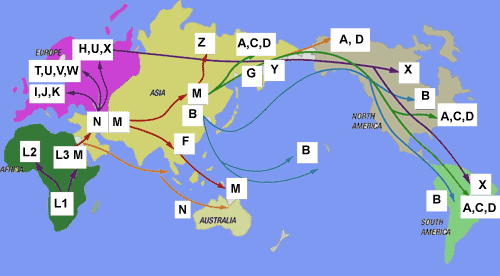
Oto wyobrażenie
pozaafrykańskiej ekspansji (migracji) ludzkości.
Rezultaty "out of Africa" i migracji: zaludnienie
kuli ziemskiej
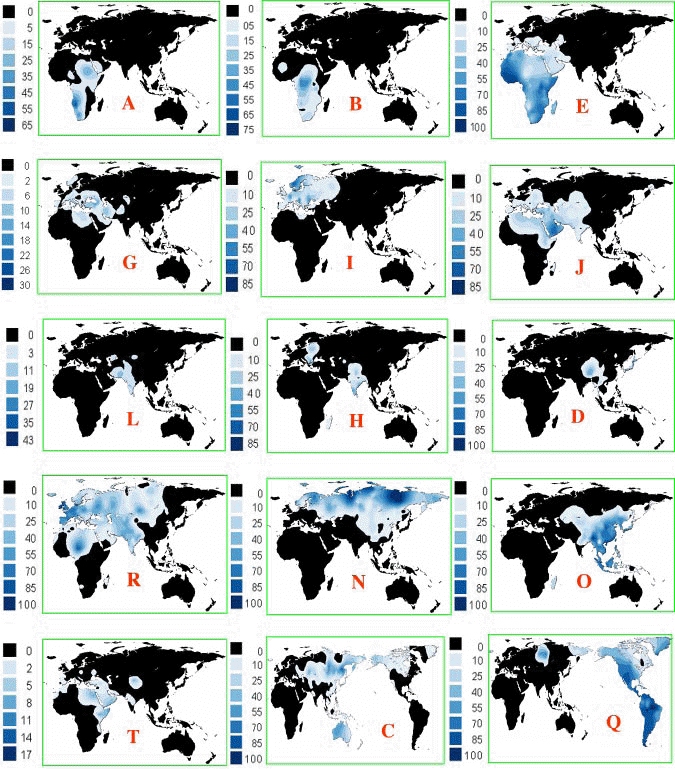
Są to mapy dzisiejszej lokalizacji poszczególnych
głównych haplogrup/rodów Y-DNA w świecie,
* * * * *
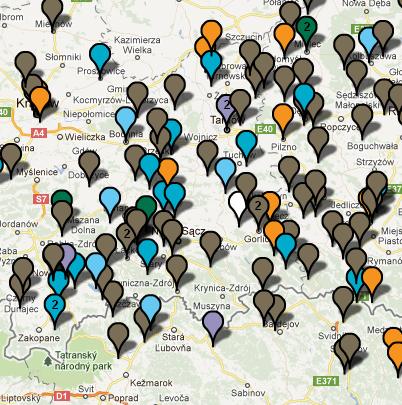
II. Haplogrupy /rody/ ojcowskie Y-DNA Z czterech ośrodków naukowych, to jest Internastional Y-DNA Project, Polish FTDNA Project , Zamagurie Region DNA Project i Carpatho-Rusyn DNA Project, udało się (na początku roku 2011) zidentyfikować około 150 testowanych Y-DNA i około 80 mtDNA z rejonu Dunajca, Popradu i Białej Tarnowskiej oraz przylegających terenów, zwłaszcza posko-słowackiego pogranicza. To już dość dużo dla wstępnego ustalenia głównych linii rodowodowych mieszkańców tego rejonu. Z Y-DNA będą tu omówione najpierw te haplogrupy/rody, które występują u nas w mniejszości; i w tej kolejności, w jakiej znajdują się one na drzewie geneal.ogicznym, idąc "od korzeni" ku górze, ku najmłodszym częściom drzewa.
1. Rody A00 A0 i A1 – pierwotne populacje afrykańskie 10 listopada 2012 r. na międzynarodowej konfernecji administratorów projektów badawczych FTDNA w Houston (USA) administrator projektu haplogrupy A, p. Bonnie Schrack, oficjalnie ogłosiła wyniki ostatnich testowań w haplogrupie A0 i wykrycie jeszcze jednej, daleko straszej haplogrupy, nazwanej A00. Jest to jakby druga połowa ludzkości, z której pierwsza, ta nasza od A do R1a, liczy około 7 miliardów ludzi, a ta druga, A00, zaledwie może kilkadziesiąt czy kilkaset tysięcy (wśród plemion Bangwa, Mbo i może Bamileke) w Południowo-Zachodnim Kamerunie. Publikacja ukazała się 28.02.2013. Wspólny przodek tych dwóch linii, pod którym nastąpiło genetyczne rozdzielenie, czyli tzw. Y-Adam, żył - według moich uściślonych obliczeń - około 250.000 lat temu (zob.). Ogólnie przyjmuje się 200 do 280 tysięcy lat. Na internetowych portalach pojawiły się, podobnie jak w kwietniu, określenia, że to "zaskakujący sukces"; "niezwykłe", "szokujące", "ekscytujące", "największe odkrycie w genetyce", "najważniejsze odkrycie w antropologii" itd., a nawet: "to odkrycie nie jest tym, które zdarza się tylko raz w czasie życia; takie zdarza się tylko raz w historii ludzkości" (This isn't just a once-in-a-lifetime event, it's a once-in-the-history-of-mankind event.). Mieszkańców rejonu Dunajca może zainteresować i ten fakt, że zarówno w prywatnej korespondencji, jak i w publicznym oświadczeniu na konferencji od p. Bonnie Schrack w Houston i w komentarzach pojawiła się informacja, że powinienem być dumny z tak wielkiego odkrycia, bo nie byłoby go, gdyby nie moja inicjatywa, ukierunkowanie i finansowe wsparcie. "Stan Pietrzak from Poland has been one of the project's most generous and enthusiastic supporters". Zob. na naszej stronie Historia odkrycia Y-Adama oraz Fundusz rozwoju rodziny A00. 2. Zespolona haplogrupa CF
a) Z
zespolonych haplogrup BT i CF, powstałych około 64000-70000 lat temu w
Afryce, wypłynęła kolejna zespolona, DE, datowana na 48000 lat.
Z niej zaś wywodzą się dwie
haplogrupy, D i E. Ta pierwsza, haplogrupa D,
powstała
zapewne już poza Afryką. Jej populacje znajdujemy dość
daleko poza Afryką, licznie w Azji Środkowej,
w Tybecie (50% ludności), Azji Południowo-Wschodniej i w Japonii
(35%). 3. Haplogrupa F
Uważa się, że zapewne ta
haplogrupa, jako pierwsza, dokonała slynnego “out of Africa”.
Opuściwszy Afrykę, około 70 000 lat temu, podzieliła się na C i
F. Najpierw wydzieliła
haplogrupę C,
która szlakiem poludniowej Azji udala się na Cejlon, do Japonii,
Melanezji, Polinezji. Jej mutacje występują wśród Aborygenów
australijskich, plemion północno-wschodniej Syberii i wśród
rdzennych Amerykańów. Nielicznie wraz z Hunami w V wieku dostała
sie do Europy. 4. Ród F – jako praojcowie pozaafrykańskiej ludzkości.
Haplogrupa F
powstała ona w zespolonej CF najpóźniej między
60000-48 000 lat temu. Za jej ojczyznę uważa się półwysep
Arabski lub Indyjski. W Indiach
jej zagęszczenie wynosi 12% ludności. Wśród bliższych nam Kurdów
7-41%. Haplogrupa F
stała się ojcowską dla haplogrupy G, H i IJK, a przez tę
ostatnią dla wszystkich pozostałych grup ludzkości (I, J, K, L,
M, N, O, P, Q, R, S i T). W sumie około 90% ludzkości! Naukowcy
W.
Haak et al. na podstawie badań kopalnego Y-DNA z Derenburgu w
Niemczech w lisotpadzie 2010 r. ogłoszają, że haplogrupy F*
razem z G2a3 brały udział we wprowadzeniu (albo przynajmniej
przyjęciu przyniesionej przez inną populację) pierwszej kultury
neolitycznej: rolnictwa i ceramiki liniowej rytej (LBK) z Azji Mniejszej do
Środkowej Europy. a) Ród G, synowska dla F, powstała w rejonie Kaukazu. Tam np. wśród ludności Gruzji i Azebejdżanu występuje w 30%. W kilku-kilkunastu procentach występuje także w niektórych krajach europejskich, zwłaszcza na Sardynii i w Szwajcarii. Populacjom haplogrupy G i F przypisuje się udział w sprowadzeniu lub przyjęciu z Bliskiego Wschodu pierwszych elementów neolitycznego rolnictwa (Derenburg, Niemcy, około 3200 p.n.e.). W Polsce rozpoznano dotąd 40 osób tej grupy, czyli 3% ludności. W naszym rejonie - 4 osoby G2a3: Uście Gorlickie, Mielec, Nowy Targ i Mszana Dolna (nie wpisane do PolishProject). b) Ród H natomiast powstała około 35 000 lat temu na Półwyspie Indyjskim i żyje tam do dziś. Nieliczne jej plemiona, jak np. Cyganie i Romowie, przywędrowali na Bałkany i przez Rumunię, Węgry i Słowację Wschodnią rozpierzchła rzadko się po pozostałej Europie. W Polsce genowo rozpoznano tylko dwie osoby haplogrupy H. W naszym rejonie tej haplogrupy genowo nie zidentyfikowano jeszcze nikogo. Ta ważna zespolona haplogrupa wydała najpierw z siebie kolejną zespoloną haplogrupę IJ. Powstała ona około 45 000 lat temu. Przez długie tysiąclecia przed Wielkim Zlodowaceniem (ok. 20000-12000 lat temu) tworzyła znane przedlodowcowe kultury archeologiczne w Europie i pozostawiła ludzkie szczątki kromaniońskie, paleoeuropoidalne. Z niej na skutek mutacji M170, także jeszcze w epoce przedlodowcowej, wyłoniła się kolejna haplogrupa /genetyczny ród/. a) Ród I, uważana więc za rdzennie europejską. Zapewne ona obok ojcowskiej IJ stanowiła w okresie przedlodowcowym w Europie jedyną populację paleoeuropoidalną. Najstarsze jej szczątki kostne i wytwory materialne zidentyfikiowano w Kostienkach nad Donem (Rosja) i datowano radiowęglowo dokładnie na 30 000 lat temu. Zapewne ona tworzyła przedlodowcowe kultury grawecką i CroMagnon. Ludność ta posługiwała się zapewne jakimiś formami języków przedindoeuropejskich, których śladów trzeba się dopatrywać w starym nazewnictwie wodnym (hydronimii) albo słownictwie niektórych języków europejskich, np. kreolskich (zlepkowych) języków germańskich. Nieco później w haplogrupie I doszło do podziału na haplogrupy I1 i I2. b) Ród I1 oraz I2b około 6000 lat przed Chr. tworzyły archeologiczne kultury protogermańskie, np. Kongemose, w rejonie Jutlandii i Południowej Skandynawii. Około roku 1000 przed Chr. obserwujemy ich istotny udział w etnogenezie Pragermanów, o czym świadczy i analiza Y-DNA archeologicznych szczątków w Lichtensteinhohle. W Polsce liczba przedstawicieli tej haplogrupy liczy dziś 8%. Rozpoznano też 4 osoby w naszym naddunajeckim rejonie: Gródek n. D, Nowy Targ, Groń i Nowa Biała na polskim Spiszu. Jest to zapewne ślad dawnego napływu na polski Spisz kolonistów niemieckich ze słowackiego Spisza, sprowadzonych tam przez władców węgierskich w XII wieku. Powstała na drodze mutacji M423. c) Ród I2a2 i jej podgrupy związane są głównie z Górami Dynarskimi (Chorwacja i Bośnia-Hercogowina), gdzie ich zagęszczenie wynosi blisko 50% ludności. Zapewne brały one udział w rozwoju niektórych archeologicznych kultur słowiańskich, np. Cucuteni-Trypolskiej.
W Polsce liczba hg I2a2 wynosi 8% ludności. Genowo zidentyfikowano u nas 7 osób (12% całej ludności) tej “słowiańskiej” haplolgrupy także w naszym rejonie: Nowy Targ, Roztoka Mała, Tęgoborze, Doły (nad Białą), Bistuszowa, Dębica i Wielopole pod Tarnowem, co wynosi 12% ludności. Z zespolonej haplogrupy IJK wywodzi się też, wyłoniona około 30 000 lat temu prawdopodobnie na Półwyspie Arabskim, haplogrupa J. Podzieliła się wnet na siostrzane haplogrupy J1 (około 20 000 lat temu) i J2. d) Ród J1 (jot jeden), wyróżnia szczególnie narody semickie: Arabów i Żydów, wśród nich znanego Patriarchę Abrahama i jego potomków. U Żydów i Arabów zagęszczenie haplogrupy J1 dochodzi do 70% (na Płw. Arabskim). W Europie Południowej bywa w zagęszczedniu od kilku do kilkunastu procent. W Polsce hg. J1 około 2,5%. Dwie osoby haplogrupy J1 zidentyfikowano także w naszym rejonie: Vyśne Rużbachy (SK) i Ciężkowice. e) Ród J2 powstała około 18000 lat temu na Bliskim Wschodzie, a największe jej zagęszczenie znajduje się w rejolnie Libanu i Syrii – około 25%. Ludnośc ta rozwijała sie w rejonie tzw. Żyznego Półksiężyca – pasa ziemi idącego łukiem Dolnej Mezopotamii, poprzez podgórze irańskiego Zagrosu, Syrię i Liban ku Delcie Nilu. Tam rodził się początek upraw zbożowych i hodowli bydła. Na terenie Syrii ludnośc ta stworzyła najstarszą kulturę ceramiczną, tzw. ceramikę impresso (‘Cardium Pottery’): pierwsze garnki z wypalanej gliny, ozdabiane odciskami muszli skorupiaków morskich, zwanych po łacinie cardium (stąd cardium pottery). Na przełomie mezolitu i neolitu, około 5500 przed Chr., pod wpływem napierajacych innych plemion ze Wschodu ludność kultury impresso ruszyła na Zachód, osiedlając się na wyspach Mora Śródziemnedgo i południowych nadmorskich krańcach Europy, od Grecji po Hiszpanię, dając im pierwsze zaczątki neolitu z ceramiką i rolnictwem. W Polsce ludności haplogrupy J2 jest ponad 5%. W naszym rejonie zidentyfikowano trzy osoby haplogrupy J2 (Łukowica, Maniowy i Słopnice), czyli podobnie 5% tego społeczeństwa. 6. Ród K Wyłoniony z zespolonej haplogrupy IJK, powstał prawdopodobnie na Półwyspie Indyjskim. Zapewne w tym regionie Azji wydzieliły się dwie kolejne haplogrupy: L (Indie, Pakistan, Sri Lanka) i T (nieliczna w Indiach i zachodniej Eurazji). Po migracji przedstawiciela rodu K w kierunku południowo-wschodniej Azji, wyłoniła się bardzo ważna grupa, którą można nazwać sundajską, K2 z mutacją M526.
7. Ród K2-M526 - Sundajski
W czerwcu 2014 T. Karafet i zespół ogłosili bardzo ważne wyniki badań genetycznych na terenie Azji Poludniowo-Wschodniej, łącznie z wyspami Indopnezji i sąsiednich krajów wyspiarskich. Tam w czasach Wielkiego Zlodowacenia (do około 13.000 lat przed Chr., gdy poziom oceanów był niższy do około 140 m, istniało duże południowe przedłużenie lądowe Półwyspu Indochińskiego, zwanego obecnie Sundaland. Tam właśnie wśród sprzyjajacych okoliczności w rodzie K2-M526 szybko rozwinęły się liczne gałęzie, dające początek licznym dzisiejszum populacjom kontynentalnym i wyspiarskim, bardzo różnym pod względem rasowym.
Kiedy około 13-10 tysięcy lat temu wielkie czasze lodowe w znacznym stopniu stopniały i wody oceanów podniosły się o ponad 100 metrów, znaczna część Sundlandu znalazła się pod wodą. Wtedy powstały wielkie, bardzo gęsto zaludnione wyspy, zwane sundajskie: Sumatra, Jawa, Bali, Timor, Celebes, Borneo i Mandanao (Filipiny). Jedne haplogrupy w całości lub prawie w całości pozostały na wyspach, żyjąc jakby w przymusowej separacji demograficznej, np. M i S oraz małe podgrupy K2. Inne haplogrupy ratowały się głównie rychłą migracją w kierunku północnym lub północno-zachodnim, np. N, O, P*, P1, Q i R. a) Ród M; powstał na Sundalandzie i dziś zaludnia niektóre wyspy Indonezji i Oceanii oraz Malaje; w niektórych rejonach zagęszcza się do 100%. Jego rasa papuasoidalna (wzrost około 150-160 cm). W Polsce nie występuje. b) Ród S - podobnie, zaludnia niektóre wyspy Indonezji i Oceanii (Papua NG); rasa papuasoidalna. c) Ród N. - powstał i żyje na niektórych wyspach sundajskich; migrował głównie w kierunku północnym. na pograniczu Chin, Mongolii i południowej Syberii. Zaludnia głównie Syberię (w Jakucji do 75%) oraz północną Rosję europejską od Uralu do Finladndii (60%) i Samów w pn. Szwecji (40%). Wyróżnia ich język ugrofiński. Z nadwołżańskich Ugrofinów wywodzili się przywódcy stepowych Madziarów (Węgów), którzy nad Dunajem utworzyli państwo węgierskie, sami w końcu ulegając tam wymarciu. W krajach nadbałtyckich haplogrupa N zagęszcza się do 45%. W połączeniu ze Słowianami tworzyła kiedyś część Bałto-Słowian. W PolishProjekt L. Mayki, obejmującym tez Litwę, występują w zagęszczeniu 7%. W naszym rejonie dotąd zidentyfikwano trzy osoby haplogrupy N (czyli 5%): w Szaryskim Jastrabiu (SK), Ostrowsku k. Nowego Targu i w Tarnowie. Zapewne to zabłądzeni tu dawni Węgrzy. d) Ród O związany jest współną mutacją a rodem N, powstał i żyje na niektórych wyspach Sundajskich, Melanezji, Polinezji i Mikronezji; migrował w kierunku północnym; jego zagęszczenie występuje gównie na terenie Indochin, Chin, Mongolii, Korei, Japonii oraz Cejlonu. 7a. Ród P, oznaczony też K2b2, z mutacją P295. Od niego pochodzi grupa P*, czyli mieszkańcy sundajskiej wysp Timor, Sumba i Sulavesi (Celebes) oraz karłowate plemię Aeta na filipińskiej wyspie Mandanao. P1 z mutacją P27 to haplogrupa, powstała na Sundalandzie pod mutacją P295 (czyli w rodzie P). Dała ona początek dwóch haplogrup: Q i R. 8. Ród Q także powstał na Sundalandzie i żyje nadal na wyspie Bali. Po migracji ród ten zaludnił znaczne obszary Syberii (międzyrzecze Obu i Jeniseju), rejon Kamczatki i Alaski oraz obrzeża Grenlandii. Haplogrupa Q stanowi podstawową część (niekiedy do 100%) rdzennej ludności Ameryki, zwłaszcza Południowej, przy niewielkim procencie haplogrupy C. Ich przejście przez Syberię, cieśninę Beringa, Kamczatkę i Alaskę do Ameryki datuje się na okolo 13 000 lat przed Chr. W Polsce haplogrupę Q znaleziono dotąd u 18 osób. W rejonie Dunajca i Popradu nie wystepuje. 9. Ród R – to głównie Europejczycy R1a i R1b
W sensie
genealogicznym haplogrupa R-M207 jest najmłodsza, zajmuje jakby
wierzchołek drzewa genealogicznego ludzkości. Dominuje też swoją
liczebnością, a także dokonaniami cywilizacyjnymi. Powstała na
drodze mutacji M207 zapewne na Sundalandzie, choć przedtem
wskazywano raczej tyllko Azję środkową. Haplogrupa R “dorobiła się” potomstwa szczególnie
ważnego dla Europy. To dwie wielkie siostrzane gałęzie:
haplogrupa R1a, która wyróżnia zwłaszcza ludy słowiańskie
oraz haplogrupa R1b, cechująca zwłaszcza ludy
zachodnioeuropejskie. 10. Ród R1b i jego podgrupy (głównie R1b1b2, Italo-Celtowie i ludy germańskie)
Haplogrupa (Ród) R1b1b2 przybyła do Europy około połowy III tysiaclecia przed Chr. Liczy ona w Europie kilkadziesiąt różnych mutacji, a więc i tyleż gałęzi drzewa filogenetycznego. Jej mutacje to bratnie P312 i U106, tworzące ważne haplogrupy (aktualnie oznaczane jako R1b1b2a1a2 i R1b1b2a1a1) i dające początek dwom wielkim populacjom. Pierwsza, R-P312, rozwiją się głównie wśród zachodnich ludów italo-celtyckich (Hiszpania, Francja, Włochy, Angia i Irlandia); przybyła tam z Bliskiego Wschodu prawdopodobnie dwoma szlakami: przez Afrykę Północną, Gibraltar i Półwysep Pirenejski (bliżej o tym zob.) lub przez Bałkany. Druga, R-U106, bratnia dla R-P312, rozwija się głównie wśród ludów germańskich i anglosaskich; przybyła do północno-zachodnich Niemiec zapewne także od Półwyspu Pirenejskiego. W Polsce z tych ważnych, zachodnioeuropejskich populacji zidentyfikowano około 16% ludności, a na naszym terenie dotąd osiem osób z tego rodu (13% całej ludności) w miejsowościach: Brzesko, Bogucice, Osobnica, Jasło, Krasne Lasocice, Łącko, Huta Wysowska i Jarabina. Podsumowując, zauważamy, że spośród powyższych genetycznych gałęzi ludzkości w rejonie Dunajca, Popradu i Białej pojawiają się nielicznie przedstawiciele tylko siedmiu z nich: E, I1 i I2, J1 i J2, N oraz zachodnioeuropejskiej R1b. Łącznie 30 osób, czyli 43% całej społeczności (bez osobnych projektów Spisza i Karpato-Rusińskiego, zob. niżej). Prawie tak samo wygląda sumaryczna statystyka całej ludności Polski. Grupy te, pierwotnie niesłowiańskie, już dawno, może przed dwoma-trzema tysiącami lat zostały zeslawizowane i zintegrowane z tymi, którzy dziś nazywają się Słowianami i Polakami. Przez kogo zeslawizowane?
* * * * * III. Genetyczny ród R1a i jego podgrupy- pionierzy indoeuropejskości i słowiańskości W pierwszej części artykułu, wyżej, wyjaśniono pojęcia genealogii genetycznej Y-DNA i mtDNA, na tle ludności świata omówiono spotykane w rejonie Dunajca haplogrupy, czyli rody ojcowskie Y-DNA, a szczególnie genetyczny ród R1b, który stanowi zasadniczą część ludności Europy Zachodniej, a naszym rejonie reprezentowany jest przez około 13 procent społeczeństwa. Drugą część artykułu poświęcimy bratniemu dla rodu R1b rodowi R1a, który również genetycznie ewoluował z haplogrupy /rodu/ R1-M173. Burzliwie rozwijająca się genealogia genetyczna i odkrycie nowych węzłowych mutacji doprowadziły do tego, że przed rokiem przygotowany do druku materiał, musiał teraz być od nowa w całości opracowany. * * * * * Mówimy tu o wielkiej populacji, liczącej kilkaset milionów ludzi, rozprzestrzenionej w w Europie i Azji od Wysp Brytyjskich do i od Uralu do Alp, z centrum zagęszczenia do 57 procent ludności w Polsce i ws niektórych okęgach Rosji; podobnie jest w naszym, naddunajeckim regionie. Wymaga więc ona specjalnego omówienia.
Badaniem genealogii rodu R1a i jego podgrup zajmują się w świecie szczególnie trzy ośrodki naukowe. 1) Od 2007 roku rosyjscy biochemicy i genealogowie Anatolij Klosow (AKlyosov, Moskwa-Boston) oraz Igor Rożanski (I. Rozhanski, Ukraina-Japonia). Dane i wyniki analiz i dyskusje publikują w portalu Rodstvo.ru, miesięczniku Вестник Российской Академии ДНК-генеалогии (Proceedings of the Russian Academy of DNA Genealogy); dostęp do tej publikacji: http://aklyosov.home.comcast.net/~aklyosov/ ) oraz w niedawno powstałym półroczniku Advances in Anthropology: http://www.scirp.org/journal/aa/ (vol 1 i 2). Podobne prace prowadzi rosyjska grupa Gentis i jej potal MolGen. 2) W ramach Polisch Project FTDNA między innymi grupą genetyczną R1a początkowo zajmowali się dwaj Amerykanie małopolskiego pochodzenia, Lawrence Mayka, prowadzący Polish FamilyTree DNA Project: http://www.familytreedna.com/public/polish/ oraz wspomagający go analityk Joseph Gwozdz: .http://www.gwozdz.org/PolishClades.html#PolishProject . 3) Natomiast ostatnio, od roku 2010, cenną inicjatywę i aktywność wykazał Łukasz Łapiński i jego międzynarodowy zespół, do którego przystąpiło wielu polskich, rozyjskich i zachodnich genealogów genetycznych. Prowadzi on międzynarodowy portal dla całej światowej gałęziu R1a: “R1a1a and Subclades Y-DNA Project”, http://www.familytreedna.com/public/R1a/default.aspx . * * * * * Haplogrupa R1a powstała w ojcowskim rodzie R1-M173 kilkanaście tysięcy lat temu. Stało się to w Azji, zapewne po odejściu R1 z rejonu Sundalandu, choć do niedawna praojczyznę R1a widziano przeważnie na terenie Iranu północnego, a wcześniej - nawet Indii.
Praojczyzna Indoeuropejczyków
Obecnie ród R1a,
z mutacją M417 i młodszymi, występuje licznie w Azji Środkowej,
np. w niektórych plemionach Kirgizji do około
70%, i w Azji Południowej (Pakistan, Iran i Indie – po kilkanaście
procent, ale to oznacza w Indiach około 150 milionów ludzi, a wśród
kapłańskiej kasty braminów nawet 72%!). Licznie występuje także w Europie Środkowej i Wschodniej. W
Rosji i Ukrainie około 45-50%, na Słowacji – około 40%,
w Czechach około 35% ludności;
dokładniej zob. w Wikipedii: http://www.en.wikipedia.org/wiki/Haplogroup_R1a_(Y-DNA)
W Polsce
natomiast ludności rodu R1a jest najwięcej spośród
krajów Europy, bo ok. 55-57% społeczeństwa.
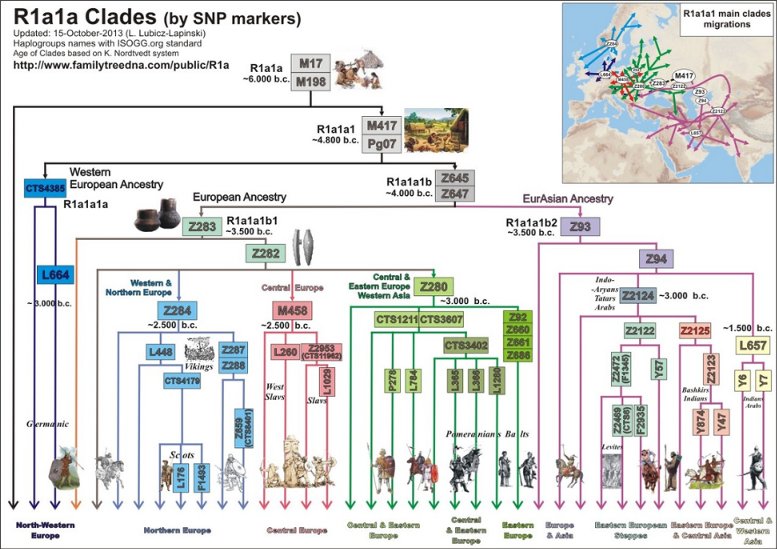
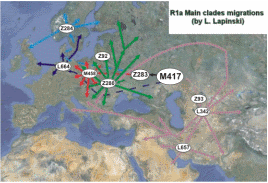
W ramach światowego programu pełnego testowania "1000 Genomów" wykryto w 2011 roku kilka węzłowych mutacji SNP w genetycznym rodzie R1a. Ułatwiają one teraz budowanie poprawnego drzewa filogenetycznego, opracowanie geografii rozprzestrzenienia poszczególnych podgrup oraz kojarzenie ich obecności i migracji z wielkimi procesami historycznymi. Oto aktualne drzewo filogenetyczne rodu R1a, opracowane przez Igora Rożanskiego.
Na powyższym diagramie nie uwzględniono jeszcze świeżo odkrytych SNP i modyfikacji tego drzewa na skutek wydajnego aktualnie, od końca 2012 r. prowadzonego projektu Geno 2.0
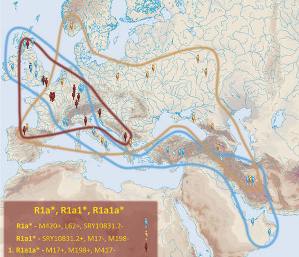
A. a) Rody R1a, R1a1, R1a1a
Mapa polska (L. Łapiński)
Źródło: http://www.familytreedna.com/public/R1a/default.aspx?section=results
Powyższe mapy należą już raczej do
przeszłości, gdyż większość owych próbek Y-DNA została, po
kolejnych testach, przyporządkowana młodszym grupom, jak niżej.
Przypuszcza się, że takie grupy powinny nadal znajdować się
raczej gdzieś w Azji, na szlaku migracji
od Sundalandu ku Europie.
B.
Patrząc ponownie na powyższy diagram drzewa filogenetycznego, na jego pniu zauważamy zapewne autochtonną, europejską mutację M417 (czerwony punkt) określoną obecnie na drzewie jako R1a1a1, powstałą około 6800 lat temu (4800 przed Chr.).
1) Mutacja
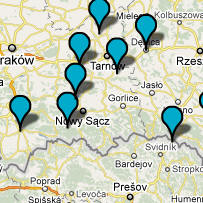
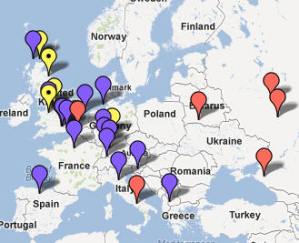
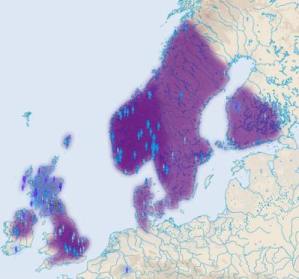
CTS4385 i jej podgrupa L664 W czasie około 3800 lat przed Chr. w rodzie M417 u jednego mężczyzny powstała mutacja CTS4385, a ona zapoczątkowała ród zwany dziesiętnikami, gdyż w markerze DYS388 wyjątkowo ma 10 alleli. Najstarsze populacje tej haplogrupy jednak wyginęły, skoro wspólny przodek obecnie żyjącej populacji żył dopiero około 2800 lat przed Chr. (zob. na mapie poniżej, pineski żółte). Ród ten, podobnie jak poprzednio omówione, znów rozprzestrzenił się prawie tylko w północno-zachodniej Europie, o czym informuje poniższa mapka. Stąd i jego nazwa: "gałąź północno-zachodnia".
Ta
północno-zachodnia haplogrupa sięga nawet na północnoszkockie wyspy
Orkady. A właśnie tam niedawno odkryto ośrodek starej i wielkiej
kultury archeologicznej, z megalitami starszymi niż piramidy
egipskie i dużym ośrodkiem świątynnym. Najstarsze z nich
powstały około 3200 lat przed Chr.
2) Mutacja Z93 Mutacja Z93 na drzewie genetycznym stanowi obecnie haplogrupę R1a1a1h. Powstała ona w rodzie z mutacją M417, zapewne gdzieś w Europie wschodniej, około 3500 lat przed Chr. Jest to wielka populacja określana ogólnie jako azjatycka, bo rozprzestrzeniła się w Azji na przestrzeni wielkiego trójkąta: od Morza Kaspijskiego i Południowego Uralu do Kirgizji na wschodzie i Indii na południu. Od początku musiała żyć niejako na uboczu pozostałej części M417, zapewne w rejonie andronowskiej kultury Sitashta (a grodem Arkaim w rejonie Południowego Uralu), skoro niezmieszana z innymi wcześniejszymi grupami tego rodu, rozproszyła się w Azji. W niektórych plemionach Kirgizji liczebność tego rodu sięga 70% ludności, a w Indiach - około 16 %, czyli co najmniej 150 milionów. I co ciekawego - w górnej kaście hinduskich braminów liczebność tej mutacji sięga aż 72 %. Lud tej mutacji zdobył te przywileje zapewne jako najeźdźcy, przybysze a Europejskiej praojczyzny. Głównie ten ród, Z93 i jego podgrupy, zwłaszcza w Indiach i Iranie, występuje w starożytnych pismach pod nazwą Ariowie. Musiała ona do nich przylgnąć jeszcze przed migracją na południe ku Indiom i Iranowi, która nastąpiła około połowy 2 tysiąclecia przed Chr., zapewne z terenu kultury andronowskiej (do tej nazwy "Ariowie" nawiązywali np. niemieccy rasiści, zmyślając mit rasy aryjskiej). Jednak Rosjanie z grupy Klosowa cały ród R1a, zwłaszcza z mutacją Z280, nazywają Ariami - zapewnie niesłusznie. Trzeba dodać, że z tej grupy aryjskiej Z93 wywodzi się też ród aszkenazyjskich żydów-lewitów, który rozprzestrzenił się zwłaszcza w pasie między Morzem Czarnym a Bałtyckim (ale Hitler o ich genetycznym pochodzeniu nie wiedział!).
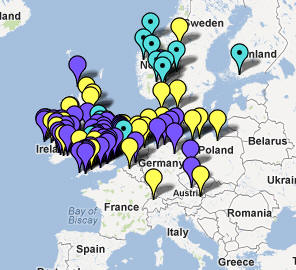
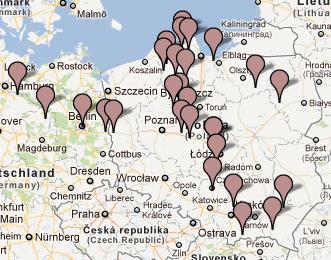
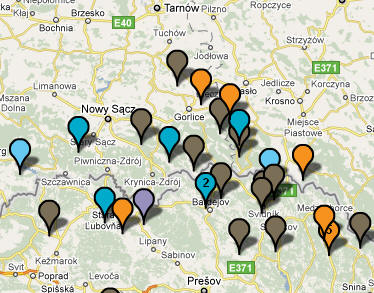
Rosyjska mapa wskazuje przedstawicieli rodu Z93
w dwóch miejscowościach naszego regionu: w Bieczu i Mielcu.
3) Mutacja Z283
Ten ród rozprzestrzeniony jest szeroko w Europie
od Alp i Łaby po Ural, a także w Skandynawii
południowo-zachodniej i stamtąd częściowo także na Wyspach Brytyjskich. Rodowi
Z283 trzeba przypisać uformowanie i rozwój ważnego zespołu
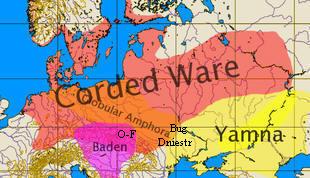
("horyzontu") archeologicznych kultur ceramiki sznurowej w
Europie Środkowej i Wschodniej oraz w
południowej Skandynawii. Kultura ta zapoczątkowana została około 3200 lat
przed Chr., a więc daleko wcześniej niż w Europie Zachodniej i
Środkowej pojawiała się kultura ceramiki dzwonowej,
przypisywana zachodnioeuropejskim rodom R1b.
Mniej więcej wtedy, około 4.000 lat przed Chr., ród Z283 podzielił się pod względem geograficznym, ale i pod względem genetycznym, według trzech zasadniczych mutacji: Z284, Z280 i M458 (zob. wyżej na drzewie filogenetycznym).
1. Skandynawski ród Z284
Głównie w tym rodzie
genealodzy chcą widzieć buszujących po
Europie wczesnośredniowiecznych rozbójników-rycerzy
i
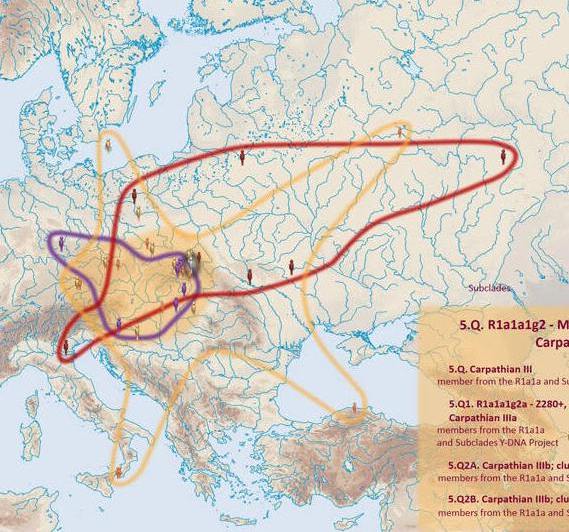
wytrawnych żeglarzy, zwanych wikingami. 2, Bałtosłowiański ród Z280 (na drzewie jako R1a1a1g2)
Drugi, również wydzielony spod mutacji Z283, ród - to wielka
grupa Z280, obecnie oznaczana na drzewie jako R1a1a1g2. Jest
szeroko rozprzestrzeniona od Łaby w Europy Środkowej do Uralu na
Wschodzie, Ród ten bywa nazywany bałtosłowiańskim, gdyż w nim i
na tym terenie kształtowały się kiedyś ludy pra słowiańskie i
prabałtyjskie Językowo była to więc grupa
jakby pośrednia między zespołem praindoeuropejskim a prasłowiańskim.
a) Gałęzie
pomorskie, kaszubskie i pruskie, z mutacjami L365 i L366
Trzech ludzi grupy L365 zidentyfikowano w naszym regionie, w miejscowościach: Łącko, Niedzica i Łapszanka. Natomiast rozmieszczona na wschód od Dolnej Wisły grupa pruska, w której niektórzy legitymują się mutacją L366, jest mniej liczna i nie ma przedstawicieli w naszym regionie. b) gałęzie północno i wschodniosłowiańskie
Kilka gałęzi północnych i wschodnich rozprzestrzenia się
pomiędzy Bałtykiem a Uralem, dość rzadko, może z powodu braku
bardziej intensywnego testowania. Niektóre ich populacje żyją jakby zanurzone w
populacjach północnowschodniej haplogrupy uralskiej i
ugrofińskiej N1a3 i zapewne uczestniczyły kiedyś, przed
tysiącami lat, w genezie Bałtów.
c) Trzy gałęzie tzw. karpackie
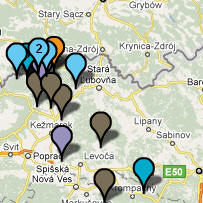
W rejonie Dunajca testowano dotąd sześć osób tej gałęzi, w miejscowościach: Tarnów, Mielec, Jabłonica i Moszczenica koło Biecza, Ciche k. Białego Dunajca i Ludomirova na Słowacji.
c-2) Gałąź
Wołga-Karpaty.
W rejonie Dunajca testowano dotąd siedem osób tej grupy, w miejscowościach: Róża w pow. Dębica, Tarnów, Łazy Biegonickie (Nowy Sącz), Gorlice, Trybsz, Białka i Frankova na Spiszu.
c-3) Gałąź
stara karpacka
YP343 z cechą DYS452=28
W rejonie Dunajca zidentyfikowano pięciu przedstawicieli
tej gałęzi, w miejscowościach:
Bartne koło Gorlic, Bystrzyca koło Dębicy, Łukowica koło
Podegrodzia (6 osób) i Matiaszowce na słowackim Spiszu.
3. Środkowoeuropejski ród
M458 (na drzewie jako R1a1a1g1)
a) Z mutacją M458 - środkowoeuropejska
W rejonie Dunajca zidentyfikowano dotąd pięciu
mieszkańców z tej gałęzi, w miejscowościach:
b) Z mutacją L260 - zachodniosłowiańska
W rejonie Dunajca testowano dotąd około dwanaście osób z
tej gałęzi, w miejscowościach:
C. (nieaktualne)
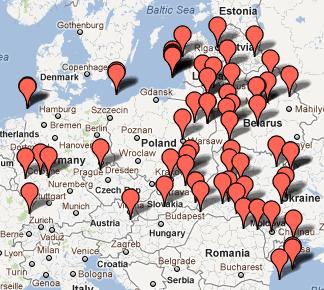
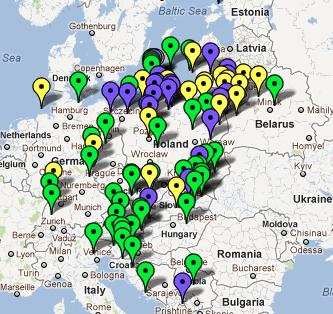
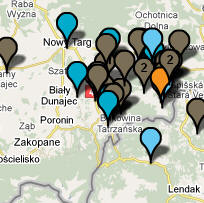
D. 1. Polski Spisz i Slovensky Spiś (z map: Zamagurie Region DNA Project FTDNA) Na Polskim Spiszu spotykamy się obecnie chyba z największym w całej Eurazji zagęszczeniem haplogrupy R1a1a1, bowiem wynosi tu ono aż 73% (odnośna mapa obejmuje 1600 km2).
2. Karpato-Rusini, w tym Łemkowie (mapa: Carpatho-Rusyn DNA Project FTDNA)
Na
powyższych dwóch mapkach niektóre haplotypy R1a1a1 są już
uwględnione na poprzednich mapkach. Ich ogólna liczba daje
jakieś wyobrażenie o procentowym udziale tego rodu w całości
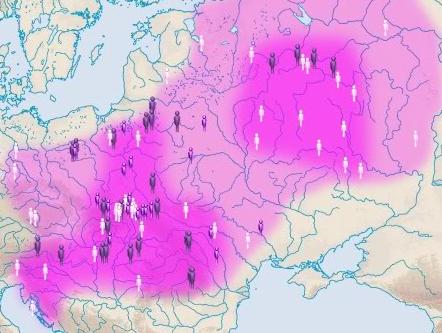
społeczeństwa. * * * * * W sumie wydaje się, że osadnictwo terenu górskiego w rejonie Tatr i na wschód od nich nadaje pewną, niewielką specyfikę ludności rejonu Dunaca. Na polskim Spiszu zauważa się np. zaskakująco podwyższony procent rodu R1a (może najwyższy w Europie), wbrew rzekomo intensywnej kolonizacji niemieckości w średniowieczu w nowotarszczyźnie i na Spiszu, zakodowanej w nazwach wielu miejscowości. Druga specyfika tego górskiego regionu to nieco wyższy procent osadników haplogrupy bałkańskiej E-V13, co może świadczyć o częściowo bałkańskim źródle migracji ludności wołoskiej i łemkowskiej, potwierdzonym badaniami matczynych genów mtDNA. Dokładniejsze badania ludności góralskiej i wołoskiej (np. Ochotnica) mogłyby dokładniej odslonić charakter ich interesujących odrębności kulturowych: czy za nimi nie kryją się także jakieś częściowe odrębności genetyczne.
Trzecia specyfika rejonu
karpackiego to zjawisko jakiegoś większego zagęszczenia liczby
gałęzi rodu R1a na omawianym terenie w porównaniu z
innymi regionami Europy środkowej i Wschodniej. Może to
świadczyć o nieco większej starodawności osadnictwa tej grupy w
rejonie Dunajca. Dlatego rosyjscy genealodzy genetyczni,
wbrew niektórym kierunkom w polskiej archeologii, nie w Rosji,
ale właśnie w rejonie Karpat upatrują regionu, z którego
dokonała się ekspansja wielu gałęzi rodu R1a w Europie, m.in. ku
Ukrainie i Rosji * * * * * IV. Język rodu R1a. Znaczenie tego rodu dla cywilizacji
1. Języki indoeuropejskie. Odkąd w 1786 r. W. Jones stwierdził wielkie podobieństwo języka hinduskiego sanskrytu i większości języków europejskich i że muszą wszystkie one pochodzić z jednego źródła, a F. Bopp w 1816 odtworzył niektóre szczegóły gramatyki owego prajęzyka indoeuropejskiego, językoznawcy nie przestają aż do dziś głowi się na temat miejsca i czasu powstania tego języka i sposobu jego rozprzestrzenienia się. Opracowano co najmniej kilkanaście teorii, z nich niektóre nadal są żywo dyskutowane. Klasyczna indoeuropeistyka wskazywała ojczyznę wspólnego języka praindoeuropejskiego przeważnie na stepach nadczarnomorskich, a koniec tej wspólnoty na trzecie tysiąclecie przed Chr. Około roku 2000 przed Chr. miał sie zacząć jej rozpad, połączony z geograficznym rozprzestrzenianiem się i dalszym różnicowaniem jego dialektów. Ostatnio wielu autorów upatruje tej ojczyzny wspólnoty języka praindoeuropejskiego na Bliskim Wschodzie, lub w Anatolii (w dzisiejszej Turcji), na Bałkanach albo wreszcie w basenie Dunaju. Natomiast czas tego rozpadu przesuwają dość daleko do tyłu, do szóstego czy ósmego tysiąclecia przed Chr. Poszukiwania nadal trwają. Ale oto mamy teraz genealogię genetyczną. Dopiero ona natknęła się na element, który kiedyś ściśle łączył co najmniej piątą część ludności półwyspu Indyjskiego i Iranu z ludnością znacznej części Europy. Jest to ród, wyróżniający się wspólnymi mutacjami SNP w Y-DNA w haplogrupie R1a1a1 – M420, SRY10831.2, M198 i M417. Ten ostatni snip istniał już około 4500 lat przed Chr. Ród, który powstał od owego przodka z mutacją M417, zapewne wówczas niewielki, musiał wnet potem rozpaść się i rozejść. Jedna część została w Europie, i w niej doszło u jednego mężczyzny do mutacji Z283. Jego potomkowie zdominowali liczebnie całą rodową populację. Druga część migrowała w kierunku Azji, za Morze Kaspijskie. Zapewne w trakcie tego migrowania u jednego z jej mężczyzn doszło do mutacji Z93 i powstania haplogrupy R1a1a1h.Wspólny przodek europejskiego Z283, i azjatyckiego Z93 żył około 3500 lat przed Chr.; do rozejścia więc musiało dojść po tym czasie. Potomstwo mężczyzny z mutacją Z93 również zdominowało liczebnie całą rodzinną populację, rozrosło się i rozprzestrzeniło w kierunku Kirgizji, Iranu i Indii. Grupa południowa, indo-irańska, gdzieś po drodze (Arkaim, Sintastha, Andronovo, dolina Amu-Darii?) wytworzyła wysoką kulturę materialną i duchową i z jej elementami wkroczyła najpierw do Indii, a nieco później do Iranu. W Indiach zawładnęła zwłaszcza kultem religijnym, tworząc tam, około roku tysięcznego przed Chr., słynne święte teksty (hymny) pod nazwą Wedy (=Wiedza). W najwyższej kaście kapłańskiej braminów liczebność europejskich "najeźdźców" sięga 72 procent. Podobnie bywa do dziś we władzach politycznych Indii, co niekiedy staje się przyczyną sporów politycznych, a nawet krwawych zajść. Podobne sukcesy osiągnęła na Wyżynie Irańskiej inna grupa rodu R1a1a1h. Jej tamtejszym dobrem jest nie tylko staroperski język z grupy indoeuropejskiej i jego nowoirańskie dialekty, ale także święte księgi Awesty.
Zarówno księgi Wedy, jak i Awesty sławią czyny i szlachetność ludu i wojowników Ariów. "Aria" ma znaczyć 'szlachetny'. Ten dumny lud mógł sam sobie nadać tę nazwę, dla odróżnienia od innych, "mniej szlachetnych" i bardziej barbarzyńskich populacji rodu R1a1a1g. Sam Dariusz, słynny władca Persji (proto-Iranu) w VI w. przed Chr., pisał o sobie "Ja, D'ariusz, syn Arii". Imię "Dariusz" też nosi w sobie etnonim "Aria". Etniczna nazwa "Iran", podobnie jak "Alan", jest odpowiednikiem etnonimu "Aria-n". Nie ma jednak racji A. Klosow, który często pisze, że nazwa "Aria" i "Ariowie" była pierwotną nazwą wszystkich populacji z rodu R1a1a1, i to już co najmniej na Rosyjskiej Równinie. Nie mają też racji dawni niemieccy rasiści, którzy całą indoeuropejską wspólnotę językową nazywali ario-germańską, przy równoczesnej sugestii, że owi Ariowie to właśnie proto-Germanie.
2. Język słowiański. Autorzy i dyskutanci w genezie etnosu słowiańskiego szczególną rolę przypisują grupie środkowoeuropejskiej (M458) i zachodnioslowiańskiej (L260), zarazem jakby odmawiając tej roli grupom północno-zachodnim, L664 i Z284, które uczestniczyły w genezie języków germańskich, oraz pomijając rolę wielkiej grupy Z280. Zupełnie niesłusznie. Przecież wszystkie te grupy pochodzą z jednego rodu, a języki tworzą się, są użytkowane i przekazywane przede wszystkim w rodzie. Wszystkie te grupy musiały więc być językowo najpierw praindoeuropejskie-satemowe, a potem prasłowiańskie, mimo że niektóre z nich, mniejszościowe, zanurzone w obcych większościowych grupach językowych, zatraciły swoją kulturową osobowość. I tak musiało się stać z grupami północno-zachodnimi. (Tu, niestety zachodzi ostry konflikt z niektórymi środowiskami archeologów, które mając zwyczaj kojarzyć języki z niektórymi formami archeologicznych kultur materialnych, wyznaczają początek języka i etnosu słowiańskiego na około V wiek po Chr., zasadniczo w rejonie Górnego Dniepru i Prypeci).
3. Języki germańskie.
4. Języki kentumowe ludów Europy Południowej i
Zachodniej.
5. Język naszego regionu. I w ten sposób, wymieniając się od stu i tysiącleci genami między sobą, jesteśmy faktycznie jedną rodziną - najpierw polską, potem słowiańską i wreszcie ogólnoludzką. Postscriptum.
Genealogia genetyczna w wielu dziedzinach naukowych, np.:
historii, archeologii, antropologii czy językoznawstwa okazuje
swoją niezwykłą przydatność do odpowiedzi na rozmaite niejako
odwieczne pytania.
I co jeszcze w tym interesujące?
*
* * *
* Dla pełnego obrazu należałoby jeszcze dodać wyniki badań DNA mitochondrialnego. W cywilizacji patriarchalnej, w której rody i plemiona tworzyły się wzdłuż męskich linni genealogicznych Y-DNA, linie matczyne (mtDNA) nie dają jasnego obrazu przynależności etnicznej. Ale mogą stanowić dodatkową informację niekiedy bardzo pomocną. W Europie i w Polsce spotykamy mitochondrialne potomstwo haplogrupy N, która (jako córka afrykańskiej haplogrupy L3) powstała zapewne na Półwyspie Arabskim po wyjściu ludności z Afryki i dała początek większości Europejczyków, podobnie jak jej genetyczna “siostra”, haplogrupa M, która dała początek większości Azjatów i autochtonnych Amerykanów.
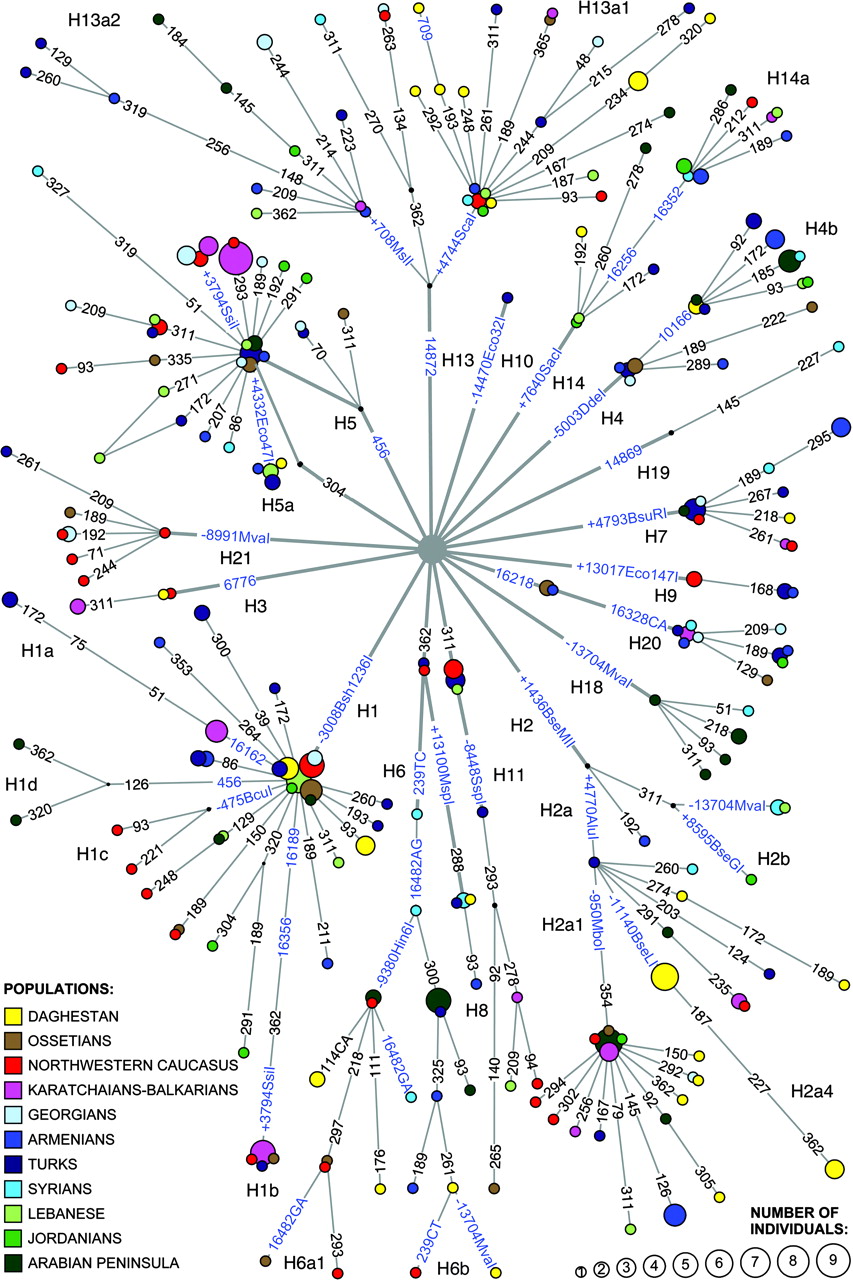
Drzewo genealogiczne haplogrup/rodów mitochondrialnych w
Europie według Eupedii:
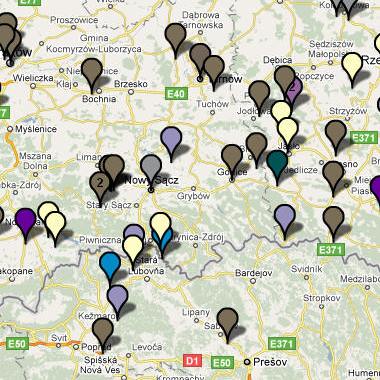
Rody mtDNA w rejonie Dunajca i przyległych okolicachPodstawowe dane lokalizacji - z map PolishProject FtDNA (niżej tablica, kolumna 1 i 2) i amerykańsko-słowackich: Spisz Polski, Spisz Słowacki i Karpato-Rusini (kolumna 3, 4 i 5)
Głównych
haplogrup /rodów/ mtDNA zidentyfikowano w świecie około czterdześci. Z
nich w rejonie
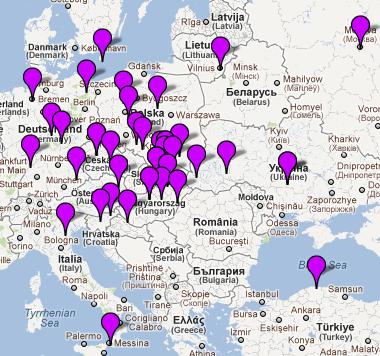
Dunajca, Popradu i Białej spotykamy osiem,
a nadto na Spiszu i wśród Karpato-Rusinów - kolejne trzy.
Szczegółowa analiza obejmie tylko dane mapy samego rejonu Dunajca. 1) Bezpośrednią “córką genetyczną" haplogrupy N jest haplogrupa W . Ta linia dotarła do Europy niezbyt licznie i zapewne z Azji Zachodniej, gdzie bywa w znacznym zagęszczeniu i gdzie zapewne powstała około 29000 lat temu. Znajdujemy tu ich przedstawicielki w dwóch miejscowościach: Toporec na Spiszu i Mały Sulin nad Popradem. 2) Drugą “córką” haplogrupy N jest haplogrupa X. X powstała około 44000 lat temu, w Europie jest niezbyt liczna i jest to jedyna grupa, która znalazła się i wśród rdzennych Amerykanów przy męskich hg. C i Q w Y-DNA, które tam się pojawiły około 15000 lat temu, przeszedłszy z Azji przez Kamczatkę, Zatokę Beringa i Alaskę. Haplogrupę X spotykamy i w Nowym Sączu (jedna osoba). 3) Bardo zasłużoną i sędziwą w Europie haplogrupą jest “wnuczka genetyczna” haplogrupy N - haplogrupa U. Towarzyszyła ona bowiem najdawniejszej ludności męskiej - w okresie przedlodowcowym, zapewne wśród męskich populacji haplogrup IJ lub I. Spotykamy ją w archeologicznych szczątkach z górnego paleolitu z czasów przedlodowcowych. Haplogrupa U3, powstała około 40000 lat temu, bardzo rzadka w Europie, najliczniej występuje w rejonie Kaukazu (około 6%), a nadto u plemion romskich Litwy, Polski i Hiszpanii (do 45%). Haplogrupa U4, powstała około 21000 lat temu, występuje częściej, rozrzucona po Europie. Haplogrupa U5, powstała 36000 lat temu, koncentruje się przede wszystkim w północnej Skandynawii i Finlandii w populacjach Saamów. Trzykrotnie spotykamy haplogrupę U i w naszych miejscowościach: Jasienna (U3), Lubicke Kupele (U4) i Litmanova (U5). 4) Kolejną jakby “prawnuczkę” haplogrupy N na naszym terenie widzimy haplogrupę HV. Najliczniej występuje na Bliskim Wschodzie, w rejonie Kaukazu i Antolii (Turcja). Liczy sobie około 17000 lat. Pojawiła się ona i w Łukowicy, świadcząc zapewne o trasie migracji dawnych plemion z Bliskiego Wschodu ku Europie Środkowej. 5) Genetyka zidentyfikowała także kilka jakby praprawnuczek haplogrupy N. Jest nią najpierw haplogrupa T, zwłaszcza T2, której powstanie ustalono na 18000 lat. Przybycie haplogrupy T2 genetyka kojarzy z przybyciem do Europy neolitycznego rolnictwa z rejonu tzw. Żyznego Półksiężyca, gdzie ta haplogrupa i zaczątki rolnictwa powstały. U nas haplogrupę T znajdujemy w dwóch miejscowościach: Toporec i Wierchomla. 6) Drugą ”praprawnuczką” jest mitochondrialna haplogrupa V. Czas powstania tej haplogrupy obliczono na 14000 lat. Była ona towarzyszką mezolitycznych myśliwych-zbieraczy. Najwieksze jej zagęszczenie dostrzeżono wśród północnoskandynawskich Saami. Jedną jej przedstawicielkę zidentyfikowano w Tarnowie. 7) Wreszcie osiem osób z naszej dwudziestki mitochondrialnej to posiadacze/posiadaczki ważnej i wielkiej w Europie haplogrupy H. Należy też do najmłodszego pokolenia, czyli “praprawnuczek” haplogrupy N. Powstała ona 18000 lat temu w rejonie Jordanii na Bliskim Wschodzie.
Stanowi ona średnio około 40% wszystkich Europejczyków, w tym i Polaków. Mitochondrialna haplogrupa H i jej podgrupy są uważane za towarzyszki dwóch wielkich europejskich haplogrup męskich Y-DNA: R1a1a1, R1b1b2 i ich podgrup, które zostały omówione wyżej. Widocznie też były ich towarzyszkami już w trakcie migracji z Azji przez Bliski Wschód i Bałkany do Europy. W naszym rejonie przedstawicielki haplogrupy H (np. H, H1e, H10, H14a) zidentyfikowano w miejscowościach: Łącko, Łukowica, Stronie, Gorlice, Wójtowa, Szynwałd, Tarnów i Radłów.
Zob. Y-DNA Inne wybrane regiony Polski
(można dzwonić
do 2100)
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



{kind=link}
.jpg){kind=link}
{kind=link}